A new paper at Nature reports on the genome of a 24,000 year-old Siberian known as Mal'ta boy or MA-1. Here's the abstract:
The origins of the First Americans remain contentious. Although Native Americans seem to be genetically most closely related to east Asians1, 2, 3, there is no consensus with regard to which specific Old World populations they are closest to 4, 5, 6, 7, 8. Here we sequence the draft genome of an approximately 24,000-year-old individual (MA-1), from Mal’ta in south-central Siberia9, to an average depth of 1×. To our knowledge this is the oldest anatomically modern human genome reported to date. The MA-1 mitochondrial genome belongs to haplogroup U, which has also been found at high frequency among Upper Palaeolithic and Mesolithic European hunter-gatherers10, 11, 12, and the Y chromosome of MA-1 is basal to modern-day western Eurasians and near the root of most Native American lineages5. Similarly, we find autosomal evidence that MA-1 is basal to modern-day western Eurasians and genetically closely related to modern-day Native Americans, with no close affinity to east Asians. This suggests that populations related to contemporary western Eurasians had a more north-easterly distribution 24,000 years ago than commonly thought. Furthermore, we estimate that 14 to 38% of Native American ancestry may originate through gene flow from this ancient population. This is likely to have occurred after the divergence of Native American ancestors from east Asian ancestors, but before the diversification of Native American populations in the New World. Gene flow from the MA-1 lineage into Native American ancestors could explain why several crania from the First Americans have been reported as bearing morphological characteristics that do not resemble those of east Asians2, 13. Sequencing of another south-central Siberian, Afontova Gora-2 dating to approximately 17,000 years ago14, revealed similar autosomal genetic signatures as MA-1, suggesting that the region was continuously occupied by humans throughout the Last Glacial Maximum. Our findings reveal that western Eurasian genetic signatures in modern-day Native Americans derive not only from post-Columbian admixture, as commonly thought, but also from a mixed ancestry of the First Americans.
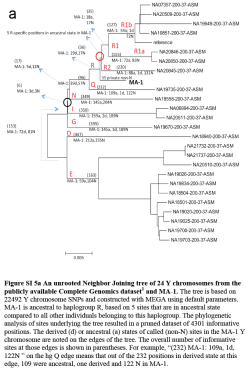
Indeed, MA-1 looks like he could be an early ancestor of present-day West Eurasians, including and especially Europeans. Mitochondrial haplogroup U was almost fixed in Upper Paleolithic and Mesolithic Europe, while R1a and R1b are, after all, the most common and widespread Y-chromosome haplogroups in Europe today.
Below is the bar graph from the K=9 ADMIXTURE analysis, which turned out to be the optimal run. Note that the Mal'ta sample appears mostly South Asian (37%), European (34%), and Amerindian (26%), but also with minor Oceanian ancestry (4%). Interestingly, among the Europeans, it's the groups from Northern and Eastern Europe that carry the highest levels of these components. This is probably a reflection, at least in large part, of their elevated indigenous European hunter-gatherer ancestry.
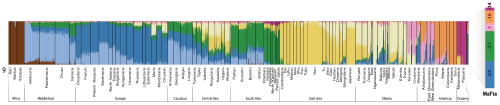
At K = 9, MA-1 is composed of five genetic components of which the two major ones make up ca. 70% of the total. The most prominent component is shown in green and is otherwise prevalent in South Asia but does also appear in the Caucasus, Near East or even Europe. The other major genetic component (dark blue) in MA-1 is the one dominant in contemporary European populations, especially among northern and northeastern Europeans. The co-presence of the European-blue and South Asian green in MA-1 can be interpreted as admixture of the two in MA-1 or, alternatively, MA-1 could represent a proto-western Eurasian prior to the split of Europeans and South Asians. This analysis cannot differentiate between these two scenarios. Most of the remaining nearly one third of the MA-1 genome is comprised of the two genetic components that make up the Native American gene pool (orange and light pink). Importantly, MA-1 completely lacks the genetic components prevalent in extant East Asians and Siberians (shown in dark and light yellow, respectively). Based on this result, it is likely that the current Siberian genetic landscape, dominated by the genetic components depicted in light and dark yellow (Figure SI 6), was formed by secondary wave(s) of immigrants from East Asia.
Here's a figure showing the levels of shared genetic drift between MA-1 and 147 present-day non-African populations. Among the Europeans it's the Lithuanians, Northwestern Russians and Baltic and Volga Finns who are most similar to the ancient sample. It's also interesting to note the relatively high position on the list of the Kalash from South Central Asia and Lezgins from the North Caucasus. At the bottom are Bedouins and Palestinians, mainly because of their non-trivial Sub-Saharan admixture, followed by Oceanians, East Asians, and South Indians, probably due to deep differentiation between their main ancestral clades and that of MA-1.

I've heard that the same team of scientists is now trying to sequence genomes from Upper Paleolithic sites west of Mal'ta. I wonder how far west? I see that the authors mention the Sungir site from near Moscow a couple of times in the paper, in relation to its similarity to the Mal'ta site. Perhaps they're working on a Sungir genome right now? If so, what's the bet that the Y-DNA turns out to be another basal R?
Citation...
Raghavan et al., Upper Palaeolithic Siberian genome reveals dual ancestry of Native Americans, Nature, (2013), Published online 20 November 2013, doi:10.1038/nature12736