I've now had a chance to look over the Lazaridis et al. preprint a few times, and also take part in several online discussions about the results, at these blogs and elsewhere. So I thought it might be useful to put together another post on the paper to report what I've learned and reiterate a few points. First of all, to understand the results, it's really important to known what the four main ancestral components in this study represent:
- West European Hunter-Gatherer (WHG), based on an 8,000 year-old genome from Loschbour, Luxembourg
- Ancient North Eurasian (ANE), based on a 24,000 year-old genome from South Siberia (dubbed Mal'ta boy or MA-1)
- Early European Farmer (EEF), based on a 7,500 year-old genome from Stuttgart, Germany, belonging to the Neolithic Linearbandkeramik (LBK) culture
- Eastern non-African (ENA), this basically means East Eurasian, and is based on samples of present-day Onge, Han Chinese and Atayal from Taiwan
Now, from what I've seen online, many people seem to think that ANE is more East Asian than European, and can be considered a signal of pretty much any population expansion from the east into Europe. This is not true. ANE is Amerindian-like, but actually also very similar to WHG. In fact, they're equidistant from ENA:
The results of Table S12.1 provide suggestive evidence that Onge share more common ancestry with hunter-gatherers than with Stuttgart. All statistics involving two hunter-gatherer populations have |Z|<0.9, so ancient Eurasian hunter-gatherers are approximately symmetrically related to Onge, and they are all more closely related to them than is Stuttgart.
We next consider the relationship of ancient samples to East Asia using the set (Ami, Atayal, Han, Naxi, She). East Asians are more closely related to all hunter-gatherers than to Stuttgart, but there are no significant differences between hunter-gatherers (all such statistics have |Z|<1.1) (Table S12.2).
...
We have conveniently labeled MA1-related ancestry “Ancient North Eurasian” because of the provenance of MA1 in Siberia, but at present we cannot be sure whether this type of ancestry originated there or was a recent migrant from some western region.
The various Uralic, Turkic and Mongolian groups expanding into Europe, usually after the Bronze Age, no doubt carried significant ENA, so these groups can't be the source of the fairly high levels of ANE across Europe today, because most Europeans lack ENA. Below is a graph based on two f4 tests, comparing ANE and ENA ancestry among Europeans, this time with the Han Chinese as ENA proxies. Note that most of the samples fall within a cline that runs from the Stuttgart sample to Estonians. The only outliers in the direction of the Han are groups from current or former Uralic and Turkic speaking areas of Europe.
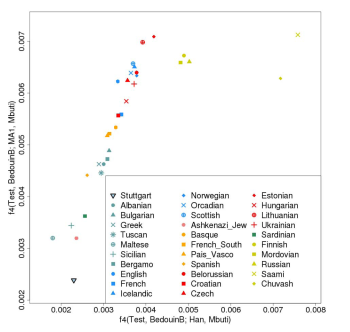
ANE was actually present in Scandinavia during the Mesolithic, because Motala12, the 8,000 year-old hunter-gatherer genome from Sweden, has an ANE ratio of 19%. But this isn't enough to explain the ANE levels carried by most present-day Europeans, so it's very likely there were at least two expansions of ANE into Europe.
Considering that Loschbour and Stuttgart totally lack ANE, it's plausible that a major wave of ANE moved across much of Europe sometime after the early Neolithic, but obviously before the Uralic and Turkic expansions, which, as per above, were rich in ENA. Based on recently published ancient mtDNA evidence from Central Europe (see here), Lazaridis et al. propose that this timeframe was the Copper and/or Bronze Age.
This of course is the generally accepted Proto-Indo-European timeframe. Indeed, the theory I put forward in the previous blog entry (see here) that most of the ANE in Europe today was the result of the Proto-Indo-European expansion, probably from Eastern Europe, looks even better on closer inspection.
Note the elongated cline formed by the European samples running from WHG to EEF on Fig 2B, shown below. It correlates well with latitude, and very likely reflects northward migrations of Neolithic farmers into Europe from the Mediterranean Basin, followed by isolation-by-distance. In other words, this cline probably took thousands of years to form.

On the other hand, there is no cline running from WHG/EEF to ANE, but all of the Indo-European and/or Eastern European samples are fairly evenly lifted up towards ANE relative to a few outliers. These outliers are all southwestern Europeans: Basques, Pais Vasco (Basque Country) Spaniards, southern French and Sardinians.
Of course, southwestern Europe is the most distant part of the continent from the generally accepted Indo-European homeland near the middle Volga. Moreover, Basques don't speak an Indo-European language, while Sardinians were only Indo-Europeanized during historic times.
Indeed, even though a couple of tables in the study report considerable ANE ancestry among Basques and Pais Vasco Spaniards, the authors admit that this need not be the case. For instance:
We next attempted to fit individual West Eurasian populations as a mixture of Loschbour and Stuttgart, as representatives of Early European farmers and West European Hunter Gatherers.
Fig. 1B suggests that this is not possible, as most Europeans form a cline that cannot be reconciled with such a mixture [Davidski's note: I think they actually mean Fig. 2B]. Nonetheless, for Sardinians (Extended Data Table 1), the most negative f3-statistic is of the form f3(Test; Loschbour, Stuttgart), which suggests that at least some Europeans may be consistent with having been formed by such a mixture. We thus fit each European population into the topology of Fig. S12.6. Only Basques, Pais_Vasco, and Sardinians, can be fit successfully with this model. Fig. S12.8 shows a successful fit.
Most European populations cannot be fit as this type of 2-way mixture and, intuitively, this is due to their tendency (Fig. 1B) towards Ancient North Eurasians that is not modeled by such a mixture.
Another intriguing thing about the results shown in Fig 2B is that the expansions of ANE across Europe appear not to have disturbed the presumably Neolithic WHG/EEF cline to any great extent. What this suggests is that ANE was spread largely independently of EEF and even WHG. In other words, the groups that pushed ANE deep into Europe probably had very high ratios of this component. This also seems to be true for the groups that brought ANE to the Near East:
A geographically parsimonious hypothesis would be that a major component of present-day European ancestry was formed in eastern Europe or western Siberia where western and eastern hunter-gatherer groups could plausibly have intermixed. Motala12 has an estimated WHG/(WHG+ANE) ratio of 81% (S12.7), higher than that estimated for the population contributing to modern Europeans (Fig. S12.14). Motala and Mal’ta are separated by 5,000km in space and about 17 thousand years in time, leaving ample room for a genetically intermediate population. The lack of WHG ancestry in the Near East (Extended Data Fig. 6, Fig. 1B) together with the presence of ANE ancestry there (Table S12.12) suggests that the population who contributed ANE ancestry there may have lacked substantial amounts of WHG ancestry, and thus have a much lower (or even zero) WHG/(WHG+ANE) ratio.
So perhaps the 17,000 year-old Afontova Gora 2 (AG2) genome from Central Siberia, classified as part of the ANE meta-population by Lazaridis et al., is genetically the closest sample we have to the Proto-Indo-Europeans? Based on a couple of the PCA from Lazaridis et al. (below) and Raghavan et al. (see here), this genome doesn't appear to be 100% ANE. My very rough estimate is 85/15 ANE/WHG.

If my assumptions are correct here, then it's no wonder that this Bronze Age Danish sample (M4) from the recent Carpenter et al. paper (see here) shows a clear shift towards the Americans on the global PCA. M4 is better known as "the old man" from the giant Borum Eshøj barrow (see here), presumably built by some of the earliest Indo-Europeans in Scandinavia. We can probably expect such Afontova Gora 2-like results from many European samples archeologically linked to the early Indo-Europeans.


{kind=link}
209 comments:
1 – 200 of 209 Newer› Newest»The problem with Finnish samples is that they don't represent the root population. The genetic difference in Fst numbers between the Finnish root population and colonial populations can be bigger than between any Central and West European populations.
It doesn't appear to be a problem though, because Finns are outliers on the ANE vs. ENA f4 graph, along with fellow Uralics (Mordovians and Sami, and even the Kargopol Russians, who surely must be significantly Uralic). So the result makes perfect sense.
It's amazing to how many groups ANE contributed ancestry --- and yet didn't survive in any unadmixed form despite living in an environment that was several million square kilometers large.
I think the main cause for the complete extinction of pure ANE groups was the low population density in their homeland, which was mainly the forest steppe. Once this zone became a highway following the domestication of the horse, it was very difficult for ANE groups to avoid admixtures both from the west and east.
But I think that if we see some ANE results of regional populations from across Russia, we might be in for a shock. Of course, none of these groups will be free of ENA admixture, but that's to be expected.
So, how early do you think ENA groups colonized siberia? It's got to be at least 15,000 yBP, but it's probably more than that.
Also their entry point: Altai?
They were there before the Ice Age (20,000-18,000 YBP), because MA-1 is 24,000 years old, and just after the Ice Age, because AG2 is 17, 000 years old. It's likely they never really left, or kept re-colonizing the area, and then mixed with the East Asians who moved in from beyond the Altai during the Mesolithic to create modern Siberians. Although most modern Siberian groups actually have fairly low ANE, except Kets, Selkups and another group that I forget the name of.
What I'm wondering is how ANE were the Andronovo groups that colonized Siberia during the Bronze Age. Their ultimate origins were in Eastern Europe, but probably an Eastern Europe very high in ANE at the time.
There's lots of sites: Sintashta, Arkaim, BMAC, etc.
No bones lying around?
Also it looks the ANE groups might have been responsible for spreading blond hair in europe since very light haired groups like Estonians, scots, and fins have high ANE ancestry. On the other hand, north caucasian groups like chechens who are also very high ANE (27%) are not light-haired at all.
Lots of remains are being tested. Check out this list from one project alone...
https://sites.google.com/site/pinhasierc/home/samples
But the problems until very recently were the technology and costs. I also have a feeling that some projects that started testing samples a couple of years ago had to go back and do it again, using more up to date methods.
That site is like...a waiting list for mummies.
No neolithic remains from southern siberia though.
I could swear though that Prof. Sarianidi was digging up whole blokes burried with horses and all though in the 80s.
It is not a problem in this study, but it is considering my target, the history of Finns. Studying the history is not a relative question, it must be based on best possible information.
I expect blond hair to be WHG connected aswell.
The European blond hair distribution largely matches the blue eyes distribution, wich again matches WHG distribution. Its just bad luck that we had a dark haired WHG in the test.
But I wonder how dark the WHG had been? Like this? ;-D
http://picload.org/image/lglprar/whggirl.jpg
btw, this is a statistik about the combination of hair and eye color in Germany:
Light blond haired Germans have:
71.2% blue eyes
12.1% mixed colored eyes
9.1% green
4.5% brown
3% grey
Middle blond haired Germans have:
51% blue
17.5% mixed
15.1% green
9.6% grey
6.8% brown
Dark blond haired Germans have:
42% blue
22.6 mixed
15.7% brown
12.9% green
6.2% grey
Light brown haired Germans have:
31.3% blue
24.7% brown
19.3% green
16% mixed
8.7% grey
Middle brown haired Germans have:
32.4% brown
25.5% blue
20.7% mixed
16.2% green
5.1% grey
Dark brown haired Germans have:
43.9% brown
22% mixed
16.9% green
14.6% blue
2.6% grey
Black haired Germans have:
63.6% brown
12.6% blue
11.9% mixed
9.3% green
2.6% grey
"Colonial populations" vs "root population" in Finland? What exactly do you mean, mikej2?
Would like to see more sampling in Volga Tatars, Crimean Tatars, Bashkirs (local R1b levels), etc. Also Balkans groups like Aromanians, Albanians, Szekely, Roma.
In Hindu Kush/Himalayas some Drokpa, Hunza, etc. More groups in India too. Small groups like this might give us some more clues.
Also better sampling in Europe should be widely used. Czechs, Polish, German regional, Swiss, Serb, Croat, Greek, etc.
It's always nice to see more samples, but I think that five well chosen ancient genomes from Eastern Europe would tell us more about the Proto-Indo-Europeans than any amount of modern samples.
Definitely. More is always better, but sampling from a variety of sites would be great. Honestly, this new paper already has more than could have been dreamed of previously.
I was also wondering if ANE shows up anywhere else (like Polynesians?).
@Davidski
"But the problem with this scenario is the tight phylogenetic relationship between ANE and WHG. If the former expanded after the LGM from a refugium in South Siberia, then why is it so closely related to the latter, which presumably recolonized Europe from a Southern European LGM refugium, basically at the opposite end of Eurasia?"
If there was a population stretching from NW Europe to Siberia before the last Ice Age then it would have been split in two by the ice and the two groups pushed towards the SW and SE respectively. After the ice retreated would you not then have two distinct but still closely related populations?
That seems the simplest explanation to me but maybe i am missing something.
#
*spaghetti
"It's amazing to how many groups ANE contributed ancestry --- and yet didn't survive in any unadmixed form despite living in an environment that was several million square kilometers large."
If they were pushed south by the ice into other people's territory and mixed there and then later re-expanded out again after the ice retreated that might be a simple explanation.
"Now, from what I've seen online, many people seem to think that ANE is more East Asian than European, and can be considered a signal of pretty much any population expansion from the east into Europe. This is not true. ANE is Amerindian-like, but actually also very similar to WHG."
I think this is a side-effect of the labeling. Eastern and Western Hyperboreans is probably not very scientific sounding.
Oy, gewalt (Facepalm). Try these reconstructions by the anatomist Gerasimov.
http://www.kunstkamera.ru/en/temporary_exhibitions/virtual/gerasimov/09/04/
http://www.kunstkamera.ru/en/temporary_exhibitions/virtual/gerasimov/09/15/
http://www.kunstkamera.ru/en/temporary_exhibitions/virtual/gerasimov/10/02/
Pirkanmiehet were Tavastians, partly from Satakunta, who moved to the Torne river valley. It is proved by Finnish historians. Karelians, Tavastians and some other people moved to Savo and populated it. Kuusamo was partly populated only 300 years ago. Proved by historians. If you want names, I can send by some private mail system. All these places were partly populated already before these movements, but the present-day population was formed by one or more root populations only a few hundred years ago. The root populations, from Southwestern Finland, Tavastia and Karelia have lived in same places much longer and can be considered Finnish root poplations from the first millennium or even before it. It is a bit same as studying Americans. They consider being Europeans, although they are present-day Americans. Europeans are the root population of white Americans. Finland-Swedes are the only enigma. Where did they come from?
NOne of these reconstructions deals with my topic:
The unusual color combination we should assume (Blue eyes and most likely blond hair on a dark skinned human) since they are all uncolored.
"If there was a population stretching from NW Europe to Siberia before the last Ice Age then it would have been split in two by the ice and the two groups pushed towards the SW and SE respectively. After the ice retreated would you not then have two distinct but still closely related populations?"
Yeah, that makes sense.
"If they were pushed south by the ice into other people's territory and mixed there and then later re-expanded out again after the ice retreated that might be a simple explanation."
But this doesn't work, because the data show that there had to be a virtually pure ANE population somewhere in Eastern Europe or Siberia during the Neolithic or even Copper Age.
"If there was a population stretching from NW Europe to Siberia before the last Ice Age then it would have been split in two by the ice and the two groups pushed towards the SW and SE respectively. After the ice retreated would you not then have two distinct but still closely related populations?"
That reminds me to the earliest theories about the split of R.
Those theories dealt with the idea that it was R1 (not a/b) that lived in Europe before the ice age and that the R1 population got turned into 2 isolated groups during the ice age (one in Spain and one in Ukraine) and that one group turned into R1a and one into R1b during the isolation....
Well, for R1b this scenario became more like unlikely meanwhile. But I could imagine it for the HGs.
There are also some less old and more western WHG reconstructions like these:
NOrwegian WHG boy:
http://www.pasthorizonspr.com/wp-content/uploads/2011/10/boy870.jpg
Swedish WHG grandmother:
http://i.imgur.com/vxu0j.jpg
And Dutch WHG grandmother:
http://skullpting.com/site/wp-content/uploads/2013/08/trijn11a-1024x713.jpg
Those are at least colored and show northern European (WHG) hair and eye pigmentation but they too, dont deal with the news that they possibly had been dark skinned.
Ah and to be complete:
The reconstruction of a farmer woman (who really does match the stereotype well (super slim face and all that)):
http://292fc373eb1b8428f75b-7f75e5eb51943043279413a54aaa858a.r38.cf3.rackcdn.com/local_05_1_temp-1368685899-51947d4b-620x348.jpg
"Well, for R1b this scenario became more like unlikely meanwhile"
Was that change (becoming more unlikely) anything to do with dating by the number of molecular mutations?
"But this doesn't work, because the data show that there had to be a virtually pure ANE population somewhere in Eastern Europe or Siberia during the Neolithic or even Copper Age."
Hmm, couldn't this be explained by layers? (This is pure speculating.) If there was an initial base population that was split in two by the ice creating the WHG and ANE halves and after some time the ice got worse such that *some* of the new ANE population in the ANE layer moved south leaving a low density unadmixed segment behind with that unadmixed segment being the first to re-expand after the ice retreated because they were both closest and best adapted to do so?
That might require a two-stage ice advance with a first advance followed by a long enough gap for WHG and ANE to develop followed by a later secondary advance that pushed some (but not all) of those ANE south. (I'll have to do some googling to see if that is plausible.)
(The same thing may have happened at the WHG end as well but if Iberia was already WHG then it wouldn't have noticed.)
Again, purely speculating but with Mal'ta boy reportedly being R* and pre ice age then the split of a base population into two halves does seem the simplest explanation (at least on the surface).
This ENA is quite vague concept, but if it is derived from Onge, Atayal and Han Chinese, it seems to be very southern entity. To me one of the biggest surprises of these Mal’ta and Afontova Gora results is the absence of East Asian and Siberian (Tungid) in pre-LGM and LGM Siberia. Of East Eurasian components, both Mal’ta and Afontova Gora possess only Native American, Arctic and Oceanian components. On the basis of these results, I would say that East Asian and Siberian components spread to Siberia only after the Ice Age or at the end of the Ice Age, and are probably connected with yDNA N and O (and some southern C clades).
It looks like pre Ice Age Siberia was dominated by yDNAs Q, R and C3, and it is these populations that made it to America.
When you refer to this big genetic difference, do you want to say that there is a big difference between Finnish and Saami populations? I agree that Finnish populations seem to have a high Baltic element (or they share a similar element/similar elements with them), and the Finnic languages were traditionally spoken around the Baltic Sea area. At the moment I am inclined to think that Saami populations came from Komi-Archangelsk areas and they assimilated pre-existing Paleo-North European populations. It also looks like there were Arctic contacts along this northernmost corridor that are reflected in the genetic composition of Saami populations. We must also remember that Finns are also said to be Finnicized Saami, as part of ancient Saamis are now part of mainstream Finns. All the more to the North we go, this assimilation of Saami populations must increase.
ENA is the Eurasian branch that includes the Tianyuan sample.
http://imageshack.com/a/img824/3230/iun5.png
It might not be too specific in the context of modern genetic substructures in Eurasia. But the important thing to understand is that it's a component shared by all present day Eastern Eurasians that isn't closely related to ANE, wasn't present in ancient Siberia, and is largely missing from modern Europe.
In other words, ANE is not ENA, and therefore it's not representative of Uralic, Turkic or Mongolian population movements into Europe.
"Was that change (becoming more unlikely) anything to do with dating by the number of molecular mutations?"
It has to do with
1. oldest R1b branches are found in Asia, not western Europe (means, R1b did branch off R1 in Asia, not in Europe), wich means, the Ice Age isolation of West and East Euros wasnt the event that created the a/b seperation in R1.
2. We continue to not find neolithic or even mesolithic R1b in Europe while the old theory had it that its even the paleolithic y-dna of western Europe.
Old (10 years) theory, some forgot already:
Paleolithic Europeans: R1a, R1b (none in the neolithic means, none in the paleolithic either)
Mesolithic migrants: I (still possible)
Neolithic migrants: E, G, J (several G and a single E have been found, so, yes)
Bronce Age migrants: N
Davidski wrote: "It doesn't appear to be a problem though, because Finns are outliers on the ANE vs. ENA f4 graph, along with fellow Uralics (Mordovians and Sami, and even the Kargopol Russians, who surely must be significantly Uralic)."
It's no wonder Northern Russians have high Siberian admixture, they Siberian component peaks in the Arctic region. It's not an ancenstral Uralic component. Siberian-ness is the highest in Norweigan Saami and forms a cline from north to south. It's a bit hard to consider it an ancestral Uralic component since it doesn't peak anywhere near the Uralic Urheimat in the Volga-Ural region.
Consensus among linguists is still that Uralic had early intimate contacts with PIE, so the two people were probably very alike and spread the same kinds of genes, if they spread any.
I don't think PIE and proto-Uralics were genetically alike at all. It's quite obvious to me that PIE were largely ANE and proto-Uralics were largely ENA. The evidence for why I think that is in my post.
"oldest R1b branches are found in Asia"
Yes, I'm wondering if the dating of the age of samples could be influenced by this:
http://dienekes.blogspot.com/2014/01/population-size-and-rate-of-evolution.html
i.e. more mutations in regions with a higher population density distorting the age estimates?
"We continue to not find neolithic or even mesolithic R1b in Europe while the old theory had it that its even the paleolithic y-dna of western Europe."
That is pretty critical obviously..
Kritiina, I have no need to speculate, we only need proper Finnish genetic samples and understanding how to create analyes.
One outcome I quite like from the paper is that it makes it quite clear why the Caucasus could appear to us to be "the womb of nations" from ADMIXTURE, even though it is not.
That's because you have a situation where the Caucasus is a fairly even, 50:50 ish, mixture of ANE and Basal Eurasian (BE), and all other populations include these components to some degree - e.g.
Northeast Euros are a mix of ANE + WHG
Atlantic Meds are a mix of BE + WHG
Southwest Asians are probably a mix of BE and southern hunter gatherers
"Gedrosians" / "Balochistanis" are a probably mixture of ANE with other complex components, possibly a differentiated, now-non existent Northern variant of Ancestral South Indian
Plus ANE is relatively closely related to WHG.
All this would tend to place a Caucasus component in a fairly central position, in terms of relatedness. But this isn't due to any Neolithic (or even paleolithic) "Caucasus race" evolving here and then having subpopulations split off. Especially combined with the mtdna evidence, which now appears somewhat misleading, probably due to high complexity in male-female population mixes (e.g. mixes might have males from one group and females from another group might then mix with males from the third group etc.).
Re: light eyes, one idea that could be possible is that hair and eye lightening alleles could have spread in the (late?) Mesolithic populations, in the Mesolithic period, through long slow selection. Then all admixed populations subsequently have the Middle Eastern origin skin lightening alleles move to fixture, after population interaction during the Neolithic, in a quick sweep.
This would explain why more ANE+WHG populations tend to be lighter eyed and haired, especially if there is some crossover where ANE+WHG light hair alleles slightly reduce skin pigment.
Think an idea of the minority blonde Melanesians who picked up Han Chinese skin lightening alleles - they might end up ulimately lighter haired and as light skinned.
Yeah, that's a good point, the West Asian womb of nations aka. Bronze Age Indo-European invasion of Europe from West Asia theory went the way of the dodo with this paper.
Mikej2, what is your opinion about the Forest-Finns in northern Värmland in Sweden? In some villages there are still people with only Finnish ancestors.
So basically the similarity of western eurasian populations isn't because they all evolved from some ur-caucasoid population, but because they have ANE, basal eurasian, and WHG components to some degree?
Yeah, it's because they share the same ancient Eurasian base and then on top of that the same or very similar recent West Eurasian components via direct and third party gene flows.
The interesting thing is that an important part of this affinity between Europe and West Asia is due to population expansions from Europe, or at least the border area in the southern Urals. This was hard to accept for a lot of people, but actually quite obvious even from basic PCAs of West Eurasia. You can see here how the North Caucasian and even Near Eastern clusters are pointing to the Volga-Ural cluster, which is probably sitting in the area of the plot where ANE came from. I don't think that's a coincidence.
http://img209.imageshack.us/img209/9898/udy9.png
Yeah, good point. And the reason Europeans are "shifted" away from west asian populations is because of WHG. And both sets also have that ANE cline which you mentioned.
But nobody ever did PCA's for native american genomes, and yet their supposed to be ANE-admixed as well. But nothing ever shows up on global admixture tests where they show up clearly as almost 100% east asian sometimes, or in their own native american cluster.
The 100% Amerindians appear to be shifted towards Europe here, including the best MA1 proxies we have, the Karitiana.
http://eurogenes.blogspot.com.au/2013/08/pca-of-world.html
Even if they don't show a shift like that on some MDS-PCA, then maybe looking at multiple dimensions might reveal something? But I'd say the shift towards Europe on the PCA above is significant.
Anna Linden: They would probably score high WHG and ANE like present day karelians. the distribution of WHG and ANE in finland is distributed so that south west finland have least of both of these components and eastern finland highest. I'm from north ostrobotnia and score more WHG and ANE than southwest finns but less than karelians because the population history show influences both from west and east finland to this region. My karelian wife score very high in both WHG and ANE.
Can someone please tell me how Davidski was able to cut a slice of K=2-K=20? I have not been able to get enough time to be able to study all the of K's if I could slice them like that it would take much less time. The study was much shorter than I excepted and to get an understanding of all it found I need to get all the info I can from the K's. They tell about nearly every population in the world.
Thank's for the answer, Mickel. My maternal grandmother was a Forest-Finn from northern Värmland. According to Anders Pålsen on "Fennoscandian Biographic Project" my Finnish genes from her are typical for people with roots in northern Karelia; but just 25 % of my ancestors are Finnish (the rest is Swedish). My WHG: 46 and ANE:18. How high is your Karelian wife's WHG and ANE? And yours?
Indeed, Karitianas are clearly shifted towards Europeans compared to other predominantly East Eurasian (ENA) populations in that PCA. But based on the same observation and PCA, most subsaharan Africans are shifted towards West Eurasians or just towards more "general" Eurasians and less so towards West Eurasians (like the Ethiopian Gumuz). Maybe we're seeing here the Basal Eurasian (y-dna haplogroup E?) and West Eurasian back-migrations... It's in line with the significant Eurasian component scored by Subsaharan Africans at lower Ks of Laziridis' ADMIXTURE run.
Anna, I know very little about Forest Finns. I have seen catalogues about them collected from Finnish sources and they seem to have been mostly from Eastern Finland, which is understandable because on the seasides Finns were not any more practiced with slash and burn method, at least not as much as people from the eastern regions. We can assume that they were mostly same people who settled wide areas in Finland during the 15th, 16th and 17th century. But speaking about genes, idk. I am a bit chagrined of this observable role of Forest Finns, because I am genealogist and from the region where many peorple moved to Sweden during hundreds years ( obviously from prehistoric to present times) and they have been forgotten totally. They were assimilated into the Swedish population. Finding them is arduous and need to go through Swedish church registers completely without help. There is in Sweden probably many times more Finnish ancestry than Forest Finns could explain. It is my estimate and I believe that it can now be seen also at 23andme's Ancestry Composition.
Me
EEF 34,69
WHG 46,61
ANE 18,70
Wife
EEF 31,00
WHG 49,22
ANE 19,78
There's a pre-print here on ancient West Eurasian admixture in southern and eastern Africa.
http://arxiv.org/abs/1307.8014
I screen capped the K=16 bar graph in several parts, and then used Gimp to stick them together. But it was a major pain in the ass, and took about 20 minutes.
I don't see what the big deal is about light eyes and darker skin complexion. Paleo hunters would have conceivably been a lot more naturalistic than modern people and less focused on display and "signaling identitykinship" (who would care out in the wilderness?).
Here are some intelligent social mammals adapted to hunting in northern environments. They are probably all brothers/sisters in this pic: http://dummidumbwit.files.wordpress.com/2010/03/wolf-pack-howling-med.jpg
Blue eyes: http://farm5.staticflickr.com/4145/5182859651_a6e807372f_z.jpg
http://ts2.mm.bing.net/th?id=H.4646563144535105&pid=1.7
The only thing I know enough about right now to comment on is the pigmentation. I can't download the study again but I do remember the ~8,000 year old Luxemburg man had "dark skin" alleles in SNP rs146554 of gene SLC24A5 but the 7,500 year old Stuggart farmer girl from Germany had the "light skin" alleles. I read from Evo and Proud that the 8,000 year old Luxemburg man did not have all three of the genes connected with European light skin, I would assume alleles of SNP's on genes TYR, SLC24A5, and SLC45A2. I also remember from the study that the ~8,000 year old Luxemburg man had the 'blue eye" alleles in SNP rs12913832, but the Stuggart farmer did not have those alleles so probably brown eyes. The Luxemburg man was said to have I think a 97.5% chance of dark hair(probably black) and the Stuggart girl had an over 98% chance of dark hair(probably dark).
I did a little research on those genes associated with certain pigmentation and distribution of pigmentation today and its connection with genetics.
The so called European light skin gene in SLC4A5 is as popular in brown skinned west Asians and some very dark south Asians as in Europeans. Meaning it does not automatically cause very pale skin like in most Europeans. It is said by Wikipedia to be 98.7 to 100% in Europeans. I trust that it does lighten skin but not in a very major way. Wikpedia says 25-40% of the difference in skin color between Europeans and west Africans is because of this gene. But is that really a big effect when considering literally black to literally white. I still think it makes a very little effect on skin color. Wikipedia and others says it is estimated to have become dominate in Europeans(which ones) 11,000-19,000ybp, a new 2014 study said 7,600-19,200ybp.. When did it become dominate in west Asians? I think it is an assumption it is only European. Just because the farmer had this gene does not mean she had light skin, she could have been very brown.
http://dienekes.blogspot.com/2013/11/europeans-and-south-asians-share-by.html
http://en.wikipedia.org/wiki/SLC24A5
TYR and SLC45A2 "light skin variants" though seem to be mainly European but still popular in west Asia and northwest Africa, and existent in south Asia. I believe the experts that say it causes paler skin, but again I doubt it makes a huge effect. TYR "light skin" variant based on that link seems to be less than 50% in Europe but SLC45A2 around 80%.
http://mbe.oxfordjournals.org/content/24/3/710/F3.expansion.html
The distribution of the 'blue eye" gene the ~8,000 year old Luxemburg man had does seem to be connected with blue eyes, but it definitely does not always mean a person has blue eyes. I trust the experts who say it causes lighter eyes, but I don't think it always causes light eyes.
Russia has over 60% light eyes and 69% of this 'blue eye' gene, Italians have 30% light eyes and 41.6% of the 'blue eye" gene, overall it seems pretty constant. According to SNpedia 80% of people with the ancestral form have brown eyes, and 99% with the derived "blue eye" form have blue eyes, so again its constant. The source I found those eye color percentages from included all light eye colors including mixed whatever that is. I know the vast majority of people with light eyes have blue eyes, but the percentage of light eyes in Europe goes down if you only count blue, so less connected with the "blue eye" gene.
Laz 2013 said there is a 52.5% chance the Luxemburg man had blue eyes, 20.7% chance of brown, and the rest mixed. There are a bunch of SNP's connected with blue eyes according to SNPedia I wonder if Luxemburg man had all of them. I think the Luxemburg man probably had blue eyes. According to a Spanish article 7,000 year old Mesolithic hunter gatherer named La Brana-1 had the same gene for blue eyes as what is in modern northern Europeans, I bet it is G,G alleles in SNP rs12913832. This is not just random, blue eyes probably existed in Mesolithic Europe.
There is really nothing to say about the Stuggart farmer she with out almost any doubt had brown eyes. Her and Otzi(early copper age farmer, ~3,250BC, Alps) both had brown eyes and are most related to modern Sardinia who have only 3.5% of the blue eye gene and probably over 95% brown eyes!!!
http://snpedia.com/index.php/Rs12913832
http://s1.zetaboards.com/anthroscape/topic/4581457/1/
http://genetiker.wordpress.com/2013/06/01/frequencies-for-blue-eyes-snp-rs12913832-in-hgdp-populations/
My best guess on pigmentation is that the ~8,000 year old Luxemburg hunter gatherer had very dark brown skin, black hair, and blue eyes and the 7,500 year old LBK farmer from Germany had brown-pale skin, black hair, and brown eyes. Otzi is reported by ancestral journeys as having fair skin, brown eyes, an brown hair. People from the Pontiac steppe from 3,000-4,000BC were reported as having pale skin and darker eyes than average modern Europeans. Bronze age iron age Indo Iranian speaking Kurgen people of Siberia and other parts of asia had pale skin and vast majority light hair and eyes. Finally the 24,000 year old Malta boy from Siberia is reported by ancestral journeys has having dark skin, dark brown hair, and brown eyes.
Today in Europe light hair, eyes, and skin are very connected which is why it is surprising the blue eyed man from Luxemburg may have had very dark skin, black hair, and blue eyes.
Here is a link with stats of eye color and hair color in European, Latin American, west Asian, and north African people. Compared with WGH ancestry, I know it is absent from the near east so far.
http://www.theapricity.com/forum/showthread.php?35882-New-Hair-and-Eye-color-statistics-(2011)
1.Sami(saw on forum one north sami person) WGH=51.57, Norway Sami light hair=49%, light eyes=68%
2.Estonian WGH=49.5, light hair=70%, light eyes=85%
3.Finn(posted results on Eurogenes blog) WGH=48, Finnish light hair=80%, light eyes=89%
4.Lithuanian WGH=46.4, light hair=57%, light eyes=78%
5. 75% Swedish, 25% Finnish(posted results on Eurogenes) WGH=46, Swedish light hair=78%, light eyes=88%, Finnish light hair=80%, light eyes=89%
6.Icelandic WGH=45.6, light hair=guess majority, light eyes: guess majority
7.Belorussian WGH=43.8, light hair=55%, light eyes=74%
8.Norweigan WGH=42.8, light hair=75%, light eyes=88%
8.Scottish WGH=42.8, light hair=52%, light eyes=80%
9.Polish(averaged two posted results from Eurogenes) WGH=41.9, light hair=46%, light eyes=68%
10.Ukrainian WGH=38.7, light hair=45%, light eyes=53%
11.Orcadien(region of Scotland) WGH=38.5, Scotland light hair=52%, light eyes=80%
12.Czech(posted results on Eurogenes) WGH=37.34, light hair=54%, light eyes=65%
13.English WGH=36.4, light hair=56%, light eyes=74%
14.Czech WGH=33.8, light hair=54%, light eyes=65%
15.French WGH=31.1, light hair=37%, light eyes=48%
16.Basque WGH=29.3, light hair=15%, light eyes=28%
17.Croatian WGH=29.3, light hair=25%, light eyes=40%
18.French south WGH=19.5, guess dark hair majority, guess brown eyes majority
19.Bergamo(in north Italy) WGH=17.7, Italy light hair=15%, light eyes=30%
20.Sardinian WGH=17.5, light hair=guess around 90%, light eyes=guess around 3-5%
21.Bulgarian WGH=14.7, light hair=10%, light eyes=27%
22.Tuscan WGH=13.6, Italy light hair=15%, light eyes=30%
23.Pais Vasco WGH=12.5, light hair guess=10-20%, light eyes guess=20-30%
24.Albanian WGH=9.5, light hair=8%, light eyes=11%
25.Spainish WGH=6.8, light hair=12%, light eyes=26%
26.Greek WGH=5.8, light hair=4%, light eyes=17%
http://www.theapricity.com/forum/showthread.php?35882-New-Hair-and-Eye-color-statistics-(2011)
http://www.theapricity.com/forum/showthread.php?107551-Y-DNA-I-found-in-Mesolithic-Sweden(-I1)-and-Luxemburg-alot-of-other-good-stuff
Blonde hair seems to have an even stronger connection to WGH than light hair. In west Europe especially Insular Celts(10-15%) red hair Even though Lithuanians had much less light hair than Estonians they had a much closer percentage of blonde hair. Blue eyes constantly took of the same percentage of light eyes. I think blue eyes are also connected to WGH ancestry(2/2 hunter gatherers probably had blue eyes).
It is crazy to say the `~8,000 year old Luxemburg man got his blue eyes(probably had them) from near eastern farmers when their closest modern relatives have very low percentages of blue eyes. Their closest Sardinia have an unbelievable low amount of blue eyes considering their WGH results of 17.5. Southern Europeans have much less WGH ancestry and much more EEF ancestry(or overall near eastern) than central, eastern, and northern Europeans they also are known for having darker skin, hair, and eyes. This is why early on I made the assumption the Mesolithic hunter gatherers of Europe were extremely pale. I except to see light hair pop up in European hunter gatherers, and pale skin(if they can somehow figure it out).
If Mesolithic European hunter gatherers had dark skin, dark hair, and blue eyes(like the Luxemburg man probably had), that means there was massive pigmentation change during the Neolithic in some Europeans or for some reason the dark hunter gatherers had few descendants.
Genetics shows there isn't that much common ancestry between all of the heavy WGH descended and pale Europeans. Something they do have in common is all of their languages and possibly most of their ancestry trace back to far eastern Europe in the copper age.
It sounds crazy but it is possible very pale skin, high amounts of light hair and eyes developed in Russia during the Neolithic, Mesolithic, or Upper Palaeolithic and rapidly spread with Indo Europeans and Uralics during the metal ages. I think it is more likely it descends from Mesolithic Europeans but maybe from eastern Europe. 2/2 blue eyes(probably) from Mesolithic west Europe probably means they also had pale skin and some light hair.
I didn't want to write too much about the pigmentation traits because everything's still so uncertain due to the low resolution data and sparse sampling from across time and space. Like I said above, let's wait and see.
Your right, I still but on blog posts not threads its okay to guess. I had no idea my posts were so long no one will read that. I still haven't studied all of Laz 2013 and cant download it anymore which sucks.
This is a shorten version so you people will actually read it.
Loschbour probably had blue eyes but maybe not, I bet he had black or just dark hair, and he could have had very dark skin or even light skin. The genes said to be European light skin genes are all about as popular in west Asia, so possibly brought in the Neolithic and there are other factors to create European pale skin Loschbour did have.
Stuggart almost definitely had brown eyes and black or just dark hair, I think brownish or light skin like modern Sardinia. Sardinia are basically LBK or related people who arrived in Sardinia in the Neolithic and almost never mixed with anyone else after that.
It is a possibility that some Mesolithic Europeans were very dark and some were very light. High amount of blue eyes in the west probably means light skin and light hair, Loschbour really doesn't make sense. Germanics and Celts probably mainly descend from Indo Europeans who spread in the metal ages, could be the same for Slavs and Balts. Finnish and Sami probably mainly descend from eastern Europeans who came there in the metal ages. The people I just named have very light skin, WGH, and high amounts of light hair and eyes. This could mean very light skin, light hair, red hair rapidly spread out of eastern Europe in the metal ages with Indo European and Uralic languages.
Maybe. What was relation between ANE and ANI?
From memory, ANI was most similar to present-day Abkhasians and Georgians, who are around 82% Near Eastern and 18% ANE. But this was a coincidence IMO, due to two or more separate migrations from the Near East and Central Asia, bringing farmers with the Neolithic package and the Indo-Aryans with R1a, respectively.
The R1a arrived no earlier than the Bronze Age, because South Asian R1a is all R1a-Z93, and thus closely related to European R1a-Z283 via the Z645 mutation. But of course the authors of that last study didn't even bother to learn about the existence of R1a-Z93, even though it was already well documented at the time.
So that Indian study will have to be done again at some point, with several well chosen ancient genomes from Eastern Europe, West Asia, Central Asia and South Asia incorporated into the models. The results and conclusions will probably look very different when that happens.
spagetiMeatball: "Also it looks the ANE groups might have been responsible for spreading blond hair.....On the other hand, north caucasian groups like chechens who are also very high ANE (27%) are not light-haired at all."
You might be right about ANE and blonde hair. Most north caucasians have more lighter skin than average Armenian or Azerbaijanian but black hair, percentage of blond hairs extremely low, but they have many reddish people. Here's some chechens
http://cs9850.vk.me/u4659473/111140455/z_4f278185.jpg
http://cs10703.vk.me/u26505840/137190654/y_146ef59b.jpg all are chechens
http://www.royor.ru/img/sport/greek-rome_12.jpg
http://www.royor.ru/img/sport/greek-rome_13.jpg
http://wsport.free.fr/2010/Aout/images/YOG/Elpiev_Anzor.jpg
http://cs320428.vk.me/v320428462/3917/_9EB9eqe94A.jpg
Among 10 chechens 2 or even 3 will be like those dudes above
Some dagestans
http://cs9589.vk.me/u16842064/118519519/z_62af038b.jpg
http://cs419625.vk.me/v419625617/257e/VuBkETPoD7Y.jpg
http://mixedarts.ru/uploads/images/00/00/16/2013/02/28/056ae9.png
In ADMIXTURE, Native Americans form their own cluster because they have a lot of drift that is specific to all Native Americans and which all Native Americans except the recently admixed tend to have.
But this isn't a guarantee that that cluster wasn't formed by ancient admixture plus drift.
ADMIXTURE makes genetic clusters which explain the data as parsimoniously as possible, even if they never fit any kind of possible phylogenetic tree.
In PCA, Native Americans often appear as only being midway-ish along the West-East Eurasia cluster, while having specific drift in a higher dimension. And sometimes there is even a West Eurasia-America dimension, with East Asians showing other drift in another dimension.
But this is hard to distinguish from actually being admixed plus having a lot of drift from just branching off early.
E.g. Oceanian people also are like the above, apparently mainly due to early branch off (although ancient genomes may tell a different story eventually).
Again, the PCA is trying for parsimony, not something that makes sense as a tree.
What ultimately helps to ascertain Native Americans as admixed is comparing how drifted they are from Africans and Europeans to how drifted an East Asian population with extra drift *should* be to Africans and Europeans.
Basically they are closer to Europeans and further from East Asians (and other ENA) than a purely East Asian + drift population should be, relative to their drift from Africans.
Initially this could appear to be due to proto-American, "East Asian" admixture in Europeans, but the drift comparisons based on the ancient Loschbaur and MA-1 genome now indiciate that that its rather because MA-1 like populations (including Loschbaur) contributed to Europeans.
Of course, historically there were suggestive results that support this. For instance the supplements in Coop et al's landmark 2009 "The Role of Geography in Human Adaptation" (which showed selective sweeps to strongly fit neutral differences) showed that the selective sweeps in Europeans that distinguish them from Palestinians are similar to the selective sweeps that distinguish Native Americans (and to a lesser extent Siberians) from East Asians. But no one has put this together until recently (and archaeological controversies backing certain theories have probably influenced this).
Most of that hair doesn't look very ginger, in a European sense, but rather reddish brown.
Some of those guys do appear kind of British, but it's hard to say if that means anything. On the other hand, that Anzor guy looks like my cousin, and could pass for a Russian too.
Anzor's chin is huge.
Don't some Algonquin-speaking native americans have(or had, in any case) chins similar to that? I've seen some old pictures and their men all looked really stern and had those robust chins.
My families from Izmir, and I've been around the meditteranean a lot and it seems to be a very rare kind of chin around there.
There needs to be some type of study that including DNA(for many reasons) to confirm these people from the north Caucasus look a certain way and their relationship to other peoples. It would be surprising if north Caucasus people who are dominated by the west Asian component looked just like central, northern, and eastern Europeans.
I thought at first maybe why they look so European is that west Asian is most related to north European which is mainly or heavily descended of WGH. But in he K's from Laz 2013 in one of the K's but I cant remember which(cant download the study anymore) there is a big purple component that dominates near east and north Africa, further down there is a big west Eurasian component, and further up near eastern components split and keep splitting and there is a very exclusively blue European component.
This would mean west Asian is really most related to southwest Asian and Meditreaen which is very surprising, because of Fst's I have seen from Eurogenes and others. Davidski says west Asian has a lot of ANE like north European which is why they seem to be related.
I don't know how else to explain the paleness in the north Caucasus, there is very little European admixture in most of them. I doubt ANE has any connection to European paleness if anything WGH does.
It is impossible to really connect red hair to WGH, EEF, or ANE because it is absent from most of Europe. Red hair only reaches over 1% in west Europe, Scandinavia(including Finnish and most Sami), parts of the Baltic, and the Urals. It may be connected to Y DNA R1b L11 which came through by eastern Europe possibly with Indo Europeans, red hair is very popular in the Urals near supposed Indo European homeland, a 3,000 year old Indo European? man from western China had a red beard, Tocharians who were Indo European had been described as red haired by Chinese and there is art depicting them with red hair(including Davidski's profile picture), A Scythian tribe in Ukraine was described as red haired by Herdotus, and Thracians who were Indo Europeans in Bulgaria were known for red hair and there is art depicting Thracians with red hair.
I think red hair may be connected with Russia, Indo European and Uralic languages. It may have been as unpopular in Europe 10,000 years ago as it was in the near east. Since red hair is so rare it is a good tool to tracing peoples origins.
I doubt red hair is at 1% in those north Caucasus people.
You can also get the study here...
http://arxiv.org/abs/1312.6639
Thank you, It worked.
Yes, but Tianyuan is not only ENA. According to Genetiker’s analyses, Tianyuan components are 35.39% Veddoid (“South_Asian”), 31.27% Sinid (“East_Asian”), 11.32% Indianid (“Amerindian”), 8.38% Nordic (“North_European”), 4.84% Tungid (“Siberian”), 3.85% Melanesid (“Australasian”),2.93% Palaenegrid (“West_African”), 1.95% Nilotid (“East_African”), 0.07% Mediterranean (“Mediterranean”). Alpine (“West_Asian”), Capoid (“Palaeo_African”), Eskimid (“Arctic”) and Orientalid (“Southwest_Asian”) are zero. Tianyuan may share c. 25% of its ancestry with ancient Siberians, i.e. Veddoid, Indianid, Nordic and Melanesid components. If we use Globe4 we get even higher figures: Tiayuan is 56.98% Mongoloid (“Asian”), 23.53% Caucasoid (“European”), 13.08% Indianid (“Amerindian”)and 6.41% Negroid (“African”). I suppose that part of Tianyuan man’s ancestry was present in ancient Siberia.
When Uralic and Turkic flows occurred, IMO these populations were already heavily ANE. No ethnic group is genetically fixed and has never been fixed. I checked the ancient mtDNAs linked with yDNA N in East Asia and found out that they vary to a big extent. When yDNAs expand, they assimilate local mtDNA wherever they go. IMO, when N arose somewhere in Yunnan&Sichuan, its carriers were autosomally Sinid. When part of them settled to Inner Mongolia/Northeast China, they became heavily Tungid, when part of them settled to South Siberia, they became heavily ANE, when part of them proceeded to the Baltic Sea area, they became mostly EEF and WHG. This logic probably applies to all populations.
To me it does not look like that Siberian mtDNA’s came from the Chinese heartland after the Ice Age. Typical Siberian mtDNAs separated from southern East Asian haplogroups before the Ice Age and many of them may ultimately have a northern origin. I propose that during the Ice Age, Northeast China, Okhotsk Sea area, Korea and Japan were important refuges for many North East Asian mtDNA lines from where they started expanding to the west at the end of the Ice Age.
I have serious doubts that Tianyuan was anything but early ENA. I don't trust analyses by any bloggers unless the results can be corroborated in some way, like by looking at the results of some samples in academic studies. How do we even know that this guy managed to call the correct variants for this genome and filter out the errors?
Also, I never said the Uralic and Turkic populations weren't heavily ANE. What I said was that they were heavily ENA, which is missing in Europe, so most Europeans don't have Uralic or Turkic ancestry, otherwise they'd have some ENA.
The ANE present in Europe had to come from another fairly late source, which basically lacked ENA, although it might well have carried some WHG and EEF.
And I have no practical experience from making analyses myself, but I have taken Geno 2 and 23andme tests. If I want to obtain my EEF, WHG and ANE percentages, may I do it myself using a calculator available online? If yes, where I could find such an on-line calculator?
How about this one? It should give a fairly reasonable result for most Northern Europeans (east and west).
http://bga101.blogspot.com.au/2013/12/eef-whg-ane-test-for-europeans.html
One posibility is yDNA Q which was found in Motala, although it was not clearly assigned to a specific haplogroup. I think that North Eurasian Q populations were originally ANE. My personal opinion is that both Eurasian Q and R were originally (mostly) ANE.
Yes, Motala12 was 19% ANE, so if that other Motala sample really belonged to Q, that would explain the admixture, because Q and R do look like ANE paternal lineages. In fact, there might have also been some R1a in Scandinavia at the time, because there's an old Northwestern European subclade of R1a, defined by the CTS4385 SNP, that is an outlier from most of the R1a found across Europe and Asia today.
But like I say above, that 19% of ANE in Mesolithic Scandinavia isn't enough to cause Northern Europeans all the way from the Atlantic to Russia to have around 15% of ANE today. So there had to be an expansion of ANE from somewhere east of Scandinavia after Motala's and Stuttgart's times, which was probably also linked to the major expansion of R1a and perhaps some R1b subclades. Interestingly, Basques actually have a lot of R1b, but are one of the few European groups that can be modeled as EEF + WHG, with no ANE.
Thank you for the link! I would like to try it with great plesure, but how can I get my Eurogenes K13 results?
You first need to upload your 23andMe data here, and then run the Eurogenes K13.
http://gedmatch.com
Actually, correction about wolves with blue eyes. Found a discussion at http://answers.yahoo.com/question/index?qid=20090320130203AAKGBxG
It turns out that wolves can have blue eyes as young cubs, but for adult wolves: "Red and Ethiopian wolves' eyes are invariably brown. Grey wolves' eyes may be brown, yellow, orange or pale green. Sometimes the pale green ones can appear blue from certain angles or in certain lights."
Although one person said white wolves can have blue eyes, which is similar to what people usually assume for humans but is contradicted by Loschbour.
My point here is that Paleo hunters would be personally bonded with the actual people in their hunting bands. By name first and foremost. They all would have seen each other as personal kin. If some were darker than others, I doubt it would have mattered.
Pigmentation in modern humans is strongly associated with ornamentation and signaling group membership. These factors would be more at play in complex high density settlements. Not so relevant to roving Paleo hunter bands IMO. If anything, hunting bands would benefit from being less picky about physical type and less xenophobic than urban/village dwellers, because hunters would benefit from new clan alliances and skills that a friendly outsider could bring into the group.
EEF farmers would have more to lose (property) by being too friendly to outsiders. They were not mobile and their stored food supplies and tools would have been a magnet for robbers and raiders. Assuming they had some kind of inheritance procedures, it would have been best to keep wealth in the family as much as possible (unfortunately leads to inbreeding depression).
Pigmentation as a visual kinship signal might have came into use with EEF farmers. Again, this could even been a way to establish a "group identity" bond/alliance between more than one originally unrelated related ancestral population. Like, say, EEF farmers (otherwise vulnerable to raids) and WHG protectors.
Yes, I meant reddish brown.Indeed, Anzor has Slavic appearance http://www.boxing-fbr.ru/uploads/photo/2010/unior/Elpiev.jpg
barakobama under red hair romans, arabian and сhinese writers meant, for instance, Anzor's hairs. Rebember Procopius of Caesarea described Slavs as red: "but they are all slightly ruddy in color", as you know Slavs aren't red. I know some guy, he's mix between Armenian (black hair, brown eyes, swarthy skin) and Ukranian (blue eyes, pale skin, dark blonde hair), and he look like this one http://www.royor.ru/img/sport/greek-rome_12.jpg Obviously north caucasians are mixed people, that's why they have more light skin.
Is there DNA evidence north Caucasians are a mixed people? From what I have seen they don't seem to be very mixed(in the modern sense), they just have some but a little bit of European admixture. DNA is the final answer in genetics you can only make guess based on appearance.
I don't know why but I have had this feeling that La Brana-1 had red hair. I had the same type of feeling or whatever I had mtDNA U5 and a subclade that had been found in Mesolithic central Europe and that is exactly what I had, and somehow I knew when I saw my great great grandfather's shotgun for the first time that my grandfather would give it to me.
I doubt the paper on La Brana-1's genome will come out soon, because they are probably using data from Laz 2013 to compare with La Brana-1. I think it is most likely he had dark hair like Luschbour. If he had red hair the researchers probably would have said so in those interviews because that would be huge news. If La Brana-1 doesn't have any of the three "light skin" genes like Luschbour that probably means there are other factors to pale skin in Europe. There is no way the combination of blue eyes and dark skin was ever common.
I know translation of those ancient writers may be rough. In the translation it says the Budni had bright red hair(seem very clear) and that there are depictions of Tocharian's that match the way Chinese writers described them.
Just took a quick check of the pigmentation genes in Loschbour. He had alleles "blue eye" alleles G,G in SNP rs1291383. According to SNPedia G,G genotype results in blue eyes 99% of the time. Laz 2013 didn't show his alleles in any other SNP's that 97% of blue eyed people have according to SNPedia.
I heard that 'blue eye" gene was used to estimate when the first person to have blue eyes lived and they estimated 6,000-10,000ybp and they connected it to the Neolithic revolution. It was said La Brana-1 had the blue eye gene(probably the same Loschbour had) those are two hunter gatherers from before 6,000 years ago, so I think that age estimate is way to recent and it wasn't in a farmer.
This link has frequencies of alleles A,A, A,G, or G,G in rs12191383 from HGDP populations.
http://genetiker.wordpress.com/2013/06/01/frequencies-for-blue-eyes-snp-rs12913832-in-hgdp-populations/
Its distribution seem close to the distribution of blue eyes, but a little to popular compared to blue eyes. I know Laz 2013 got their 52.7% probability blue eyes, 26.8% probability intermediate, and 20.7% probability brown from somewhere. I remember reading a study on the eye color in the Baltic where they used the same wording and it was through DNA but I can't find it. I think they would say that about a modern person if all they could go off of was rs12913832.
Just based on the fact Loschbour had G,G alleles I think he probably had blue eyes.
Originally everyone had A,A alleles in rs12913832 and brown eyes(unless there was another mutation that caused blue eyes). Today many areas of Europe have majority blue eyes. Not surprisingly mainly the same Europeans who scored the highest WGH. The only Europeans to score over 40% are, North Sami individual(Norwegian Sami 50% blue eyes), Estonians(69% blue eyes), Finns(72% blue eyes), Lithuanian(61% blue eyes), Icelandic(guessing around 72% like Scandinavians), Belorussian(55% blue eyes), Norwegian(72% blue eyes), Scottish(63% blue eyes), Polish(50% blue eyes).
So all so far found over 40% WGH have 50% or more blue eyes, and 2/2 Mesolithic hunter gatherers have the "blue eye" gene.
The 97.5% dark hair probability in Loschbour and 73.4% probability of black hair is unlike all of the over 40% WGH who have majority light hair some even have majority blonde hair, I am sure black hair is rare but I haven't found any statistics on it. Loschbour missing all the three 'light skin" genes that are so popular in Europe and west Asia is extremely extremely surprising. The Sami though who may have the highest WGH also have much higher amount of dark hair and eyes than their Finnish and Norse southern neighbors. In certain regions of Mesolithic Europe everyone may have had black or brown hair and other regions it was mainly light hair, just today the light haired ones have more descendants. Since no one today have vast majority Mesolithic blood I think it is possible many features became popular in the Neolithic.
barakobama: "Is there DNA evidence north Caucasians are a mixed people?"
http://en.wikipedia.org/wiki/Y-DNA_haplogroups_by_populations_of_the_Caucasus
"I think it is possible many features became popular in the Neolithic."
It's quite possible. We need more data from ancient Eastern Europeans (for instance, from Swiderian Culture or Butovo Culture or Upper Volga Culture)
Also interesting their mtDNA. According to Nasidze, chechens, avars (the biggest ethnic group in Dagestan) close to the Slavs http://onlinelibrary.wiley.com/doi/10.1046/j.1529-8817.2004.00092.x/full
Brandt showed Corded + Unetice mtDNA were like Caucasus. People assume Caucasus are "mixed" as in a mix of modern Slavs and Arabs (totally anachronistic).
But really Slavs and Caucasus might be each separately mixed of ANE + x and ANE + y. We don't know what x and y were and whether the mix was Y DNA, mito, etc.
I can't read that study now but I really doubt there are obvious similarties between the mtDNA of Slavs(which ones) and Nazidze, Checens, and Avars. I know that Caucasus people never reach 40% in mtDNA H and have somewhat different subclade percentages than any Europeans, they have a low amount of mtDNA U5 and U2e(two of the maybe the most ancient lineages in Europe), and I am guessing their J, T, and K subclades are more like near easterns than any Europeans who's subclades are basically the same as early European farmers. Trying to find relation between two west Eurasian people with mtDNA is very tricky. People are already making big claims about relation between certain modern people and ancient mtDNA, for example Bell beaker being like Iberians and LBk being like modern near easterns. Autosomal DNA is the best thing to look at, and Caucasus people always come out overwhelmingly west Asian.
From my observation some 60% of the North Caucasians have dark hair, pale skin and greenish eyes, while 30% have mostly brown, some red and some blondish hair, and another 10% dark hair darker skin (for North Caucasian standards) and dark eyes combination.
@Barakobama I assume the West Asian component is basically 70% Basal Eurasian (farmer like) + 30% ANE. While North European component is something like 50% Basal Eurasian + 50% ANE and WHG. The mixture of both components are very similar just the proportions of the mixtures are different and that West Asian or Caucasus_Gedrosia lacks WHG.
I thought the same. I assume Caucasus_Gedrosia is basically ~70% Basal Eurasian(which is farmer like I assume) and 30% ANE while North Europeans are 50% Basal Eurasians and 50% WHG+ANE. What differs these two is the missing WHG in Caucasus_Gedrosia and the different proportions of Basal Eurasian and ANE ancestry.
I haven't looked at the stats closely, but the North European component in most tests actually shows a very good correlation wth WHG/ANE, although some Europeans actually show higher WHG/ANE. What this suggests is that the North European component is purely hunter-gatherer, and on some occasions at least, some of it must even represent the WHG within EEF.
This means that the close Fst distance between the North European and West Asian components is down to the ANE within the West Asian component, which must be considerable, and perhaps as much as 40%. This correlates with PCA results, which show that Caucasian and Near Eastern groups with high levels of the West Asian component are shifted north towards Europe across the first dimension.
So Caucasus_Gedrosia is 40% ANE. Wow now thats huge but makes sense.
I had also this impression that Near Eastern populations with high West Asian shift towards East and North Europeans while populations with high Mediterranean/Southwest Asian shifted towards South Europe.
I'm not really sure how much exactly, but there's a huge difference between the more Levantine and East Mediterranean centered components and the Caucasus and Hindu Kush centered ones. That difference has to be ANE, because Northern Europeans mostly carry WHG/ANE, and even the EEF they carry is likely a hybrid component with considerable WHG.
This is clearly reflected on West Eurasian PCA where Caucasians, Iranians, Armenians, and even more northerly Semitic groups are significantly shifted north and northeast compared to many of the Saudis and Bedouins, who are almost purely Southwest Asian.
So the estimate of a 40% ANE contribution to the West Asian, Caucasus, Gedrosia and similar components might actually be an understatement. It makes sense though, because it seems there were multiple ANE expansions into West and South Central Asia, which we can see via Y-chromosome haplogroups like R1a, R1b, R2 and Q.
David there's something about autosomal DNA I don't quite get. You're saying the WHG group and the ANE group are closely related according to the study. So how can one group be from haplogroup I and the other from haplogroup R, when those paternal lineages split so much farther back?
The ANE component was found in an individual of 20 000 years of age This isn't much farther back. I shares ancestry with the macro Haplogroup K to which R belong to.
WHG estimation of EEF component is consider something along 33-2%. It is so hard to estimate the exact percentage because the Bedouins taken as Near Eastern signal in Stuttgart have considerable recent East African admixture. So the more we take this admixture into account the less EEF comes out as WHG admixed. I would go by something around the 15% range of WHG ancestry in EEF.
I agree that allot of Haplogroups in the Near East correlate with ANE ancestry and I would even add yDNA J, L and probably T to it as well.
Considering that the majority of the macro Haplogroup IJKLT belong to ANE ancestry, I doubt that J wasn't ANE connected as well if we take into account that it's closest cousin I is WHG.
I believe that especially Haplogroup G2a and sometimes E1b1b* played the major role in spreading farmer component.
What is southwest asian anyways, "basal eurasian"?
WHG and ANE look like the same population that got split into western and eastern sub-populations by the Ice Age, and then linked up again soon after. It seems that at the western end they belonged exclusively to hg I, and at the eastern end to R and Q. I suppose that in the middle, around the Urals, they might have been I, R and Q.
It's even more difficult to guess what the story is with the other haplogroups until we see more ancient genomes.
In large part yes, but without any ancient genomes from the Middle East, it's very difficult to say any more than that. We don't even know yet what Basal Eurasian is exactly.
What about west Asian in the Caucasus? Did you forget they are overwhelmingly west Eurasian? I doubt the basal Eurasian thing is accurate. There are basic physical traits in west Eurasians, if Stuttgart was 44% west Eurasian I doubt she would have had west Eurasian features.
Davidski in figure 2(p. 17) ANE is shown as a brother group to west Eurasian, and Stuttgart's west Eurasian ancestry being a brother clade to WHG. It seems to me they say west Eurasia and ANE are two separate groups, but long lost relatives. I have noticed MA1 clusters closest to west Eurasians, in K=4 he is mainly west Eurasian buts looks about 30% east Eurasian, later in the K's almost all that east Eurasian becomes native American.
I am still not entirely convinced MA1 wasn't a mutt. Y DNA R*(R1-, R2-)and mtDNA U*(distinct branch maybe should be called U-MA1), what? That doesn't seem pure to me, there was probably mixing at some point. Doesn't it make sense to say early West Eurasians around 40,000 years ago had 100% or mainly Y DNA F and IJ? WHG is really European, just modern Europeans are mainly a mix between WHG and EEF(mainly derived from the same source near easterns descend from). Loschbour was 100% European(who knows maybe 99.978%) his mtDNA was U5b1a(two 31,155ybp pre U5 samples in Europe) he had Y DNA I2a1b*(or intermediate between I2a1b and I2a1 aka pre I2a1b) doesn't it make sense his paternal lineage is upper Palaeolithic European. It is no surprise Y DNA J the brother clade to I is so popular in near easterns, because WGH and near eastern are brothers and there are many mtDNA U and RO lineages native to the near east.
Y DNA R is descended from K-M526 which is also the father to Y DNA haplogroups that are exclusive and popular in east Asians and Oceania. It makes the most sense it was originally in eastern non Africans.
ANE (MA1's relatives) contributed to the modern Amerindian and Siberian gene pools, so it's the Amerindians and Siberians who are mixed. Loschbour's relatives were always stuck in Europe, blocked in by ANE to the east, that's why there's no WHG outside of Europe, and Loschbour comes out 100% European.
Stuttgart is apparently around 44% Basal Eurasian, and the rest Middle Eastern and European West Eurasian.
There is definitely WHG outside of Europe. It doesn't matter if Upper Palaeolithic and Mesolithic Europeans never mixed with outsiders their descendants who had some EEF ancestry did. Didn't an Assyrian from the Eurogenes project get something like 4% WHG. I will have to read what the study says about MA1 to learn if he was very mixed or pure.
I am very surprised that there was almost no mixing in Europe for some 20,000 or 30,000 years. One major group of people(by genetics) dominated probably all of modern Europe. I am sure it is more complicated than that ancient mtDNA has proven U5 is descended of some of the earliest Europeans, same with U2(not U2e, 30,000ybp U2 was another lineage), possibly U4, and some rare U* lineages. RO found in Upper Palaeolithic Europe and debated RO's or descendants of RO. Then Y DNA I is very exclusive to Europe and estimated to be over 20,000 years old. Even the autosomal DNA of these hunter gatherers proves they had pure ancestry(except Motola12) from their own branch on the west Eurasian family tree.
There's definitely no WHG outside of Europe. The study says this very clearly. People who aren't of European origin aren't supposed to use the EEF/WHG/ANE test. If they're from the Near East they can use the Near East/ANE test instead.
And MA1 is not admixed. How can an individual from 24,000 years ago be a mixture of recently formed populations? Consider these quotes again from the study that I posted in the article above:
"The results of Table S12.1 provide suggestive evidence that Onge share more common ancestry with hunter-gatherers than with Stuttgart. All statistics involving two hunter-gatherer populations have |Z|<0.9, so ancient Eurasian hunter-gatherers are approximately symmetrically related to Onge, and they are all more closely related to them than is Stuttgart.
We next consider the relationship of ancient samples to East Asia using the set (Ami, Atayal, Han, Naxi, She). East Asians are more closely related to all hunter-gatherers than to Stuttgart, but there are no significant differences between hunter-gatherers (all such statistics have |Z|<1.1) (Table S12.2)."
Davidski,
I am wondering about something here. Perhaps you can correct me if this is wrong. Of course haplogroup I is a hunter-gatherer. Should not J, his brother, also be a hunter gatherer? I am wondering if perhaps J was in the Balkans or in Anatolia at this time. I am wondering if perhaps WHG was also in Anatolia or the Near East and this is where the mixing took place for EEF, as the mix is uniform across Mediterranean Europe (all 4 Neolithic Samples from Sweden to Spain and Austria). It doesn't really follow a cline from Near East to Spain like addition flow in Greeks and South Italians. I have seen some Anatolians be able to take the EEF/ANE/WHG and not go over the 100% mark on EEF, with minor WHG. Also, if there is no WHG in the Near East, why is there haplogroup I in the Near East reaching up to 10% in some places? Perhaps WHG was in the Near East at somepoint, and the Basal backflowed into the Near East from South Asia, pushing out mostly or completely absorbing WHG except in small enclaves of Anatolia. What do you think?
Davidski,
This deal with some Near Easterners having more ANE than Europeans is a bit strange. If it is Indo Europeans that pushed up the numbers and spread, why is it so more popular in some Near East populations. Could the Indo Iranians be the pure ANE, inflating the numbers there, and Indo Europeans being a mix of ANE-WHG, and possibly minor EEF in Ukraine to Kazakhstan. Perhaps a 50-50 mix for Indo Europeans as Northern Caucasus people run around 25% ANE these days.
Sorry, for the third post. It will be my last tonight. Perhaps a good chunk of this EEF in northern Europe was brought in my Indo Europeans, which is why we see so many NW Europeans getting all of these Adygei scores... just another though
I don't think hg I is native to Europe, but it obviously got there very early. Basically, I think it came from the population described as West Eurasian in this study, from which WHG broke off at some point and moved to Europe. HG J came from the same West Eurasian population, but it stayed in the Near East much longer, and probably only expanded into Europe during the metal ages, although I could well be wrong there.
In any case, it looks like modern Near Eastern populations formed when Near Eastern West Eurasians first mixed with the Basal Eurasians (or perhaps in fact Northeast Africans) during the Neolithic, and then with Indo-European and Caucasian (maybe Hurrian, Kura–Araxes, etc.) ANE coming down from the north and east.
Why wouldn't there be European hunter gatherer ancestry outside of Europe, in for example Indo Iranians and northwest Africans? MA1 is closer to us than he is to the begging of eastern non Africans and western non Africans. What I am saying is that his Y DNA R and mtDNA U is evidence he had western non African and eastern non African ancestry. He lived in Siberia very close to where eastern non Africans where at that time it is very likely east and west were mixing at that time. Everyone is admixed in someway even an Australian aboriginal.
So the hunters(including ANE hunters?) are more related to east Eurasians(takes to much room to say it the other way) than Stuttgart was? What does this mean, they had east Eurasian ancestry Stuttgart didn't have?
Only some Near Easterners have higher ANE than Indo-European speaking Europeans, and these are in fact North Caucasians. This makes sense because the North Caucasus is directly south of the regions formerly occupied by the Sredni-Stog and Khvalynsk cultures, which apparently were pre-Proto-Indo-European cultures, perhaps with almost fixed levels of ANE. But once these people expanded, they probably very quickly picked up WHG in the west, EEF in the south and ENA in the east. If so, this mostly happened via maternal gene flow, because these were almost certainly very patriarchal and patrilineal societies, which means they might have been spreading most of the same Y-chromosome lineages thousands of years after the Proto-Indo-European expansion.
But, having said all that, I have a feeling that ancient South Asian admixture in parts of the Caucasus and Near East is indirectly pushing up the ANE levels there. See the link below for some background on that...
http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0073682
I posed this question to the authors at bioRxiv, because as far as I can see, their model doesn't take into account South Asian ancestry, so it'll be interesting to see how they react to it.
Maybe we need aDNA from the earliest farming sites in the levant and anatolia. Too bad its so hard to go there, without rebels trying to shoot you or each other.
The problem with this idea is that all the Near Eastern-like ancestry across most of Europe can be explained by EEF, and this explanation actually matches the spread of early Neolithic archeology across the continent.
So Northwestern Europeans most likely show affinity to the Adygei and other similar groups via third party ANE and EEF gene flow into Europe and the North Caucasus, rather than any direct links.
Post-Neolithic gene flows from the Near East only seem to have affected Southern Europe, and this is why some of the groups there are outliers from the main European farmer/hunter cline of diversity. Perhaps this is also why Greeks have such high ANE, but barely any WHG, and as per above, maybe that's how hg J made it to Europe? This blog entry might be of interest in that respect...
http://eurogenes.blogspot.com.au/2013/06/recent-gene-flow-from-africa-and-near.html
Note the low levels of Near Eastern IBD in Northwestern Europe, which surely would be much higher if there was gene flow from West Asia into Northwestern Europe after the Neolithic.
Neolithic and Iron Age remains from Turkey, Israel and Syria have already been tested. Only mtDNA was analyzed, but it won't take long before we see some complete ancient genomes from there. The problem to date was the crappy technology.
It took some time for this to find it's way into my mind (I guess that reflects rather badly on my mental agility), but if the "Basal Eurasian" construct is pointing towards something "real", West Asians have a substantial share of ancestry that is much more divergent from West Eurasians than the ASI ancestry found in South Asia! ASI ancestry is ENA, which is much closer to West-North Eurasians than the highly divergent "Basal Eurasian" branch. This might explain why Europeans and Pakistanis-NW Indians cluster together in "trees" used to display genetic distance, to the exclusion of Southwest Asians (Bedouin, Palestinians, and the Druze). The same effect occurs using ChromoPainter/FineStructure. I often wondered why Europeans and South-Central Asians were being linked together, and separated from Southwest Asians, in these kinds of analyses. I guess "Basal Eurasian" ancestry might explain this, since ASI is less divergent from West Eurasian ancestry. But all of this is contingent on "Basal Eurasian" ancestry being a "real" factor, and not just an abstraction (although, at the end of the day, everything in historical population genetics is an abstraction).
Good observation. I'd say this is caused by a component that has greater affinity to Sub-Saharan Africa than other Eurasian components do, and the presence of this component at greater levels among some Middle Easterners than other Eurasians. The component in question might be the mysterious Basal Eurasian, or perhaps Sub-Saharan admixture, both recent and the more cryptic ancient kind, or all of the above. In fact, the Basal Eurasian component might well just be cryptic Sub-Saharan, particularly East African admixture, which is something that is considered in the paper.
"[Basal Eurasian] could reflect continuing more recent gene flows between the Near East and nearby Africa after the initial out-of-Africa dispersal, perhaps associated with the spread of Y-chromosome haplogroup E subclades from eastern Africa 9, 10 into the Near East, which appeared at least 7,000 years ago into Neolithic Europe 11."
That actually makes quite a lot of sense, perhaps "Basal Eurasian" is just a composite of different Sub-Saharan gene-flows into the Near East. But I'm having some trouble understanding the place of West Asians in the model the authors proposed. Would it be correct to say that they think West Asians are Basal Eurasian+ANE? Does WHG have any equivalent in the Near East?
Yes, it seems that EEF is only around 44% Basal Eurasian, but mostly Near Eastern. If so, this suggests the contribution to the makeup of EEF of a WHG-like West Eurasian component native to the Near East, and perhaps ancestral to WHG, as well as WHG-like component from somewhere in Europe. But the paper doesn't explicitly say that, as far as I can see, so I got it from looking at Fig. 2A and reading between the lines.
Whether that's the case or not, modern West Asians are certainly Near Eastern and ANE. In other words, they're EEF minus WHG, but with ANE.
Interesting, so there might be a West Eurasian component specific/unique to the Near East, and closely related to WHG. I hope they develop some of these angles before the paper gets formerly published.
What I find astounding, about all of this, is the amount of reticulation we've found using ancient genomic data. The past just seems to be getting more and more complex, and the closer we look, the more rhizomatic our genetic ancestry starts looking. And this is just going by ancient DNA from the Palaearctic. I wonder what awaits us if they ever do succeed in extracting ancient DNA from the tropics.
@Seinundzeit: Looking at Figure S12.19, their model for the Near Eastern populations, they have the ancient Near East being a mixture of basal Eurasian and a Loschbaur like "Western Eurasian" type population, Y, that is closest to Loschbaur under their model.
However, they haven't actually given drift statistics, so this topology could actually accomodate "Y" being no more related to Loschbaur than ANE (assume drift length West Eurasian->W is 0 and West Eurasian->Y is high).
Their model assume binary splits, so they would have to model a three way split of the ANE-WHG-"Y" clade by having "Y" share a clade with one of the other two, but no actual shared drift for this clade.
Anyway, they don't seem to model the ancient Near East as a pure basal Eurasian+ANE mix in any event, but as requiring some third WHG/ANE type element involved in some capacity.
On another note, but still related to the idea of Basal Eurasian, one thing I found questionable is how they estimated drift in both basal Eurasian and the Western Hunter Gatherers who are ancestral to Stuttgart.
Basal Eurasian itself has low levels of drift from Africans and other Eurasians, but high levels of drift are added in the WHG ancestry of Stuttgart, presumably to model Stuttgarts high levels of drift from Africans and other Eurasians.
I don't know what method they used to derive these figures. The other drift and mixture figures are constrained by having to produce Stuttgart, Loschbaur, MA-1 and Karitiana as outcomes, but for the populations that only lead to Stuttgart the only constraint seems to be that Stuttgart has to be sufficiently drifted from Loschbaur and ANE.
This doesn't seem to constrain whether drift takes place in Stuttgart's hunter gatherer ancestors after they split from Loschbaur or the basal Eurasian ancestors after splitting from the rest of the Eurasian clade. This doesn't really matter for their model, but would be important if we were looking at Basal Eurasian as a "real" population.
Any insights from your read Davidski?
My mother's second cousin's results. His ancestors were mainly Forest-Finns from northern Värmland in Sweden.
EEF: 34,5658
WHG: 46,699468
ANE: 18,734733
@Seinundzeit, Nick Patterson was kind enough to post some f_3 stats for South Asians in the comments section of Dienekes blog. They are:
## Sources Target f_3 Z
LBK Onge Vishwabrahmin 0.000734 0.641
LBK Onge Gujarati1 -0.006114 -4.582
LBK Onge Gujarati2 -0.006442 -4.955
LBK Onge Gujarati3 -0.001871 -1.325
LBK Onge Gujarati4 0.002510 1.814
MA1 Onge Vishwabrahmin 0.005237 4.136
MA1 Onge Gujarati1 0.002404 1.641
MA1 Onge Gujarati2 0.003153 2.124
MA1 Onge Gujarati3 0.004297 2.923
MA1 Onge Gujarati4 0.009156 6.084
LBK She Uzbek -0.024589 -26.829
MA1 She Uzbek -0.013565 -13.372
(There are 4 Gujarati groups in the Human Origins data, genetically distinct).
Source: comments section of http://dienekes.blogspot.com/2013/12/europeans-neolithic-farmers-mesolithic.html
If I read these outputs correctly: Some Gujaratis but not Vishwabrahmins have EEF (LBK) ancestry. Uzbeks have EEF and ANE ancestry.
Question remains whether EEF f_3 stats represent Basal admixture or actual EEF admixture from Europe. Or did EEF even originate in Europe in the first place, or just migrate to Europe from somewhere else?
Wow, exciting!
I haven't looked at why Laz 2013 thinks basal Eurasian was in Stuttgart, but I still really doubt it. I guess possibly mtDNA N1 could be connected with it and maybe Y DNA G. I think they need to consider the evidence against this in physical features. It is hard to explain that any west Eurasians have significant non west Eurasian ancestry. You can't argue that Basal Eurasian helped create the Caucasian features, because we know it didn't exist in pre Neolithic Europe and there are many examples of Caucasian skull shapes from pre Neolithic Europe. I bet the basal Eurasian thing will be proven incorrect pretty soon, it honestly sounds crazy.
Davidski, proto Indo Europeans were farmers so they would have had EEF or any type of near eastern ancestry because of that. If they were in eastern Europe they would have had a lot of WHG before they expanded. Natural human society by definition is patriarchal, and early Indo Europeans were not unique at all in that way. I do think that their societies were more aggressive and war like than most which is the main reason they spread their languages and why their Y DNA is so dominate all over the place.
R1b M269 in Europe probably originally came from the near east. If any form of it was spread by Indo Europeans they had some type of paternal connection to the near east. Y DNA R1a1a1 M417 is really the only candidate of Y DNA spread by Indo Europeans with a pure east European(WHG and ANE) or Siberian source(ANE maybe some ENA).
The whole idea is that EEF is mainly descended of farmers who arrived in Europe from the near east and mixed a little with the native hunter gatherers. Near easterns when put into EEF, WHG, and ANE admixture always come out close to 100% EEF because it is most related to what is in the near east today. OF course EEF did not originate in Europe.
There was definitely post Neolithic near eastern gene flow into the Balkans and Italy. In so many admixture's they have more west Asian or southwest Asian than north European. I have seen in PCA's they are in-between Europe and the near east around where European Jews are. Sardinians are almost completely pure descendants of Neolithic Europeans, they come out majority Meditreaen they don't have the huge amounts of west Asian and southwest Asians Italians and Balkan people have.
I think the Balkans and Italy got their near eastern ancestry from the same source. It is highest in southern Italy and Greece. At first I thought it happened during Greco Roman times. Y DNA J in Europe is highest in Italy and the Balkans, but subclade J2b is very exclusive to the Balkans and south Asia I think it has a Neolithic origin.
I think people need to start looking at Celts(Iberia-, most of France-) and Germans in western Europe as mainly descended of Indo Europeans that came from east Europe about 5,000 years ago. Are Germans very similar to Stuttgart, are the Norse like Gok4, are people in the Alps like Otzi? The Neolithic people of west Europe where probably all very Otzi or Gok4 like. That changed obviously in the metal ages because of Indo Europeans. I think the same might be true for Balts and Slavs. Once the Rise project is done, people will probably start realizing this.
I think Scandinavians including Sami mainly descend from people that came in the metal ages. I have heard some say that the reason north European(not all are in north Europe like Swiss and Ukrainians) have more WHG is because farming spread there later. I think Gok4 is prove that no matter where the farmers spread they could keep basically the same genetic makeup.
So, here are my results:
EEF 33,11
WHG 48,05
ANE 18,84
Also Sein, on this topic - West Asians have a substantial share of ancestry that is much more divergent from West Eurasians than the ASI ancestry found in South Asia! ASI ancestry is ENA, which is much closer to West-North Eurasians than the highly divergent "Basal Eurasian" branch. This might explain why Europeans and Pakistanis-NW Indians cluster together in "trees" used to display genetic distance, to the exclusion of Southwest Asians (Bedouin, Palestinians, and the Druze).
Adding up the average distances in Figure S12.12 (range of parameters), the distance between
Western_Eurasian and ENA (Eastern Non African) is
((33+43)/2)+((23+24)/2)+((32+36)/2)) = 95.5
Western_Eurasian and Basal Eurasian is
((33+43)/2)+((23+24)/2)+((23+32)/2)+(11+34)/2) = 111.5
If ASI has a drift from ENA of around 25, then ASI is further from Western_Eurasian than Basal_Eurasian is (95.5+25 = 120.5).
Onge has a drift of around 80-84 and Z (basal East Asian) 28-35 in this model, so it is certainly plausible ASI is further from Western_Eurasian than Basal_Eurasian is.
Being in a clade with another population doesn't necessarily mean those population are actually closer together than one is to an outgroup. For instance Onge to Western Eurasian divergence is around 177.5, compared to Western Eurasian to Basal Eurasian divergence of 111.5, despite WE-Onge forming a clade to the exclusion of BE. For another instance, Western Eurasian to Z (basal East Asian minus drift / admixture specific to Americans and Far Easterners) is 127.
Similarly in a phylogenetic context, if Europeans form a clade with Pakistanis that doesn't mean they're necessarily closer to them compared to South West Asians (they could be or could not).
@barakobama
I think this paper goes to show that Indo Europeans were not from the Near East. Otherwise SW Europe would probably have never gotten ANE in the amount that they do. ANE is throughout the Mediterranean, likely because of the Indo-Europeans. Not just globs of more EEF. Obviously, R1a never changed the genetics of Iberia or the rest of the Mediterranean. It's negligible in NW Europe as well. R1b had to be as much ANE or close to that of R1a for people like the Scots to be around the Lithuanians in ANE, even with R1b possibly travelling twice as far.
@About Time - Thank you very much! Makes one think, I wonder how a population closer to my neck of the woods will look.
@Matt - Thanks, that's rather interesting. So genetic distance is wholly separate from constituting a clade. I wasn't really aware of that.
R1b-M269 that is
Assyrians score even negative for WHG (the once I have seen). Iranians and Kurds usual score some 4%. I think you mixed them up.
Actually, the theory goes that the Proto-Indo-Europeans were mostly pastoralists.
I agree that they probably were partly EEF, but their EEF level couldn't have been very high. I'm not sure if I made much sense above trying to explain why, so I'll put it another way.
The ratios of EEF/WHG across Europe vary mostly with latitude and Neolithic archeology. They don't show any obvious signs of being disturbed by post-Neolithic EEF gene flows, except in Southeastern Europe.
On the other hand, ANE doesn't correlate with anything all that well, except the Indo-Europeans, because it's missing or almost missing among the non-Indo-Europeans and late Indo-Europeans of southwestern Europe, like Sardinians, French and Spanish Basques and Southwest French. But it is clearly present among all other Europeans including the Spanish.
So ANE likely arrived deep in Europe very late, like after the Neolithic, and the populations that brought it didn't seem to have a great impact on the EEF/WHG ratios already present across the continent at the time. So I think it's pretty clear that these populations had extremely high ANE ratios.
Also, another interesting thing suggested by the data is that ANE moved through Europe very rapidly, because the ANE levels in the west aren't much lower than those in the east. So the populations spreading ANE didn't mix much along the way as they were moving west. They only seem to have done that after they settled down.
WHG and ANE is much higher in Celts and Germans than in Stuttgart, Otzi, and Funnel beaker farmers, and EEF is much lower. Obviously after the Neolithic there was a huge rise of WHG. There is already plenty of DNA(not just mtDNA) from bronze and iron age Indo Iranians, the Rise project will give some from proto Balto Slavs, maybe this year there will be a lot of info from copper age Yamna people. We already know Indo Iranians probably had a lot of WHG because they had very high amounts of mtDNA U5a, U4, and U2e and would have had some EEF because they had typically near eastern farmer mtDNA haplogroups. I am sure the Corded ware and Yamna people will have a mix of near eastern farmer and European hunter gatherer mtDNA as well. They probably had WHG, ANE, and EEF. Not all of the early Indo Europeans 6,000 years ago would have been the same anyways.
There are Uralic's with high amounts of ANE and so did Motola12. It is not purely Indo European. Also I do think many ethnic groups in Europe today mainly descend from people who migrated there during the metal ages not Neolithic. When i say that I mean Celts, Germans, Slavs, Balts, Finno-Urgics, Italians, pretty much everyone but Sardinians.
One of the main reasons why WHG is much higher among Germanics and Celts than Stuttgart, Oetzi and the Funnel Beaker farmers is because it took a long time for farmers and hunter-gatherers in Western and Central Europe to mix.
It's likely that the western Indo-Europeans did bring Eastern European WHG to the west with them, but this isn't the only source of WHG in Western and Central Europe.
And I have doubts that the Indo-Iranians were significantly WHG, since WHG is missing from Asia. I'd say they were mostly ANE, with some EEF and WHG.
Also, Uralics can't be the main source of ANE in any part of Europe that isn't or wasn't Uralic speaking, because Uralics and former Uralics carry ENA, which is missing from most of Europe. Motala12 and his relatives might have passed on their ANE to modern Europeans, but his ANE ratio isn't high enough to explain most ANE ratios across Europe today.
What hunter gatherers where left in west and central Europe 5,000 years ago? I am sure very few. I know about that mtDNA U5 stuff from Neolithic Germany and the Gotland hunter gatherers. Otzi is just over 5,000 years old but was very similar to Stuttgart who is 7,500 years old. There is no evidence of a gradual rise of WHG. Gok4 was all the way in Sweden yet had around the same amount of hunter gatherer ancestry as Stuttgart. There were not nearly enough hunter gatherers in west Europe to create even close to the amount of WHG Celts and Germans have.
I will have to organize more mtDNA from FTDNA to see if modern west Europeans have mtDNA more similar to eastern or western hunter gatherers. I have heard U5b2 is mainly in central-west Europe obviously Mesolithic descended. I think the amount of WHG native to west Europe is the same amount Stuttgart had at the most.
I never said Uralics are the source of any ANE. I think Indo Europeans are not the only source. Obviously ANE in Finns is not from Indo Europeans.
How do you explain north European in the middle east especially in Indo Iranians. They had over 30% mtDNA U5, U4, and U2e. They had a pigmentation very close to Balts and Scandinavians today who have the highest amount of WHG. I think they had a lot of WHG probably over 50%.
Was there ever a credible claim of Indo-Europeans in the Near East in prehistory?
The Sumerians and their neighbors were not Indo-Europeans.
The historic instances are (1) a brief Hittite expansion into the North Levant to about Lebanon at the peak of their empire, (2) the presence of the Mittani contemporaneously at the eastern fringe of the Tigris-Eurphrates Valley, (3) te Phillistines in the Southern Levant during Bronze Age Collapse, (4) Roman rule of the Levant and Egypt, and (5) Indo-European incursions during the Crusades and European colonial era.
Nothing major, like what happened in Europe. That's for sure. I've always thought the kurgan hypothesis made more sense than anything in Anatolia.
Hunter-gatherer DNA bounced back in Europe during the Late Neolithic and Bronze Age for two reasons:
- hunter-gatherers who were hiding from farmers finally mixed with them, learned to farm, and bred at the same pace, creating hybrid settlements
- hunter-gatherers in Eastern Europe mixed with some of the farmers there, became pastoralists, invaded the rest of Europe, and Indo-Europeanized the continent
Here's some background on the first point...
http://www.researchgate.net/publication/257648697_2000_Years_of_Parallel_Societies_in_Stone_Age_Central_Europe/file/9c960525abd628afa5.pdf
Also, I doubt that most of the ANE among Finns comes from Uralics, because it's unlikely that the Uralics from the southern Urals totally replaced the earlier Indo-Europeans near the Baltic when they moved there. But we need ancient DNA from early Finns to settle this issue.
And the North European component you see in many parts of Asia most likely represents ANE, perhaps with minor WHG. Note that MA1 comes out significantly North European in ADMIXTURE analyses, and ANE is very similar to WHG anyway, so why shouldn't it be interpreted, at least partly, as North European ancestry?
We both agree WHG and ANE raised very much in west Europe and Scandinavia starting around 5,000 years ago, right? I doubt if hunter gatherers existed throughout most of west Europe during the Neolithic that there were enough to make a major impact on all the farmers. I don't understand why they suddenly began to mix rapidly during the late Neolithic. There is no evidence of that in Otzi. I guess it is possible but I think it is more likely Indo Europeans are the main reason.
You need a lot more evidence than that(Blätterhöhle Neolithic mtDNA), to say there were a bunch of hunter gatherers "hiding from farmers" in Europe during the Neolithic. Why would they hide from farmers? Why wouldn't they mix in a major way after living by each other for thousands of years? Why didn't they mix culturally? Is there any archeological evidence for this?
It seems to me the farmers in some way conquered most of Europe. There is little difference between Stuttgart an early farmer in central Europe to Gok4 a farmer nearly 2,000 years later in Sweden, and with modern Sardinians about 7,500 years later.
I have to look at mtDNA in modern Celts and Germans to make an opinion on whether most of their U5, U4, and U2e types have a Mesolithic or Indo European origin. If what your saying is true there should be some evidence in modern mtDNA.
I doubt Kunda and Comb Cermic cultures were Uralics because their mtDNA is typical hunter gatherer unlike modern Uralics. But did they come to the Baltics after Indo Europeans? Even if they did indo Europeans where never Finland.
Are ANE and WHG really that related?
I'm guessing the migrant farmers thought the hunter-gatherers were basically a bunch of apes living in the forest, while the hunter-gatherers didn't know what to make of the farmers, but they were spooked enough to stay out of their way as much as possible. That's why they didn't mix for a long time, and probably didn't have much to do with each other at all until they eventually started trading, and some hunter-gatherer women were accepted into the farmer societies when there was a shortage of females there. But once the ice was broken, they mixed very quickly, which is probably partly why hunter-gatherer mtDNA shot up in frequency among the remains from the late Neolithic.
And yeah, WHG and ANE are tight man. They're like long lost brothers.
But, having said all that, I have a feeling that ancient South Asian admixture in parts of the Caucasus and Near East is indirectly pushing up the ANE levels there. See the link below for some background on that...
http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0073682
I posed this question to the authors at bioRxiv, because as far as I can see, their model doesn't take into account South Asian ancestry, so it'll be interesting to see how they react to it.
The ANE ancestry in West Asia and the Caucasus seems to be ANI-related (thus Caucasoid), but not ASI-related, as there is virtually no ASI ancestry in West Asia and the Caucasus except in Iran and Kurds.
Also, the suggested genetic link between ancient Syria and South Asia did not persist and the South Asian-specific mtDNA haplogroups were almost completely removed from the Syrian gene pool during the subsequent periods.
Onur, you're like the lone ranger. You appear out of nowhere all guns blazing whenever someone mentions possible South Asian admixture in the Near East.
He's Turkish (as I am).... and naturally we become hectic when somebody mentions some kind of admixture into Turks whether african or indian or whatever.
According to some Kemalist authors, Turks are the pure descendants of the original seljuk nomads who entered anatolia circa 1,200 years ago.
We get touchy about this subject.
This is the source of thousands of flame wars between various assortments of greeks, turks, bulgarians, FYROMs, armenians, etc. and sometimes serbians and other balkaners join in. At the end you are confused and cannot remember who ****ed whose mother or who was the goat herding savage or who sleeps in a yurt.
It's all very entertaining.
There needs to be a mechanism for turning the steppe HGs into steppe herders.
I think the most likely is that a population of sheep-herders from eastern Anatolia wandered up to the edge of the steppe, settled by a river, mixed with the local HGs and the composite gradually developed into the Indo-European steppe culture.
So the language and some of the DNA from Anatolia, most of the DNA from the northern branch of hyperboreans and the steppe culture evolving from the environment itself.
@barak
"I doubt if hunter gatherers existed throughout most of west Europe during the Neolithic that there were enough to make a major impact on all the farmers."
Not as HGs no but if they transformed into herders then their population could have greatly increased.
.
@Davidski
"Hunter-gatherer DNA bounced back in Europe during the Late Neolithic and Bronze Age for two reasons:"
Three reasons imo: Funnelbeakers being the third.
"hunter-gatherers in Eastern Europe mixed with some of the farmers there, became pastoralists"
I think this happened along the Atlantic coast also - probably initially at the sites of valuable resources like gold mines, amber trade etc where the farmers had small colonies - leading to a "Bantu Expansion" type event from the Atlantic coast West to East i.e. Funnelbeakers. This was followed by the Indo-Europeans from the east later.
Funnelbeaker + Unetice -> Corded Ware
Funnelbeaker expansion 3000 BC (5000 BP)
http://www.geocities.ws/reginheim/funnelbeakerculture.gif
Lactose tolerance
http://img534.imageshack.us/img534/6959/lactose.png
Corded Ware
http://i129.photobucket.com/albums/p217/dpwes/cw_map.jpg
The South Asian mtDNA was from Tell Ashara (ancient Terqa) and Tell Masaikh (ancient Kar-Assurnasirpal). Dated to the period between 2500 BC and 500 AD.
Assimilated Mitannians who went back to old homelands. By that time they spoke another language. Edict of Ashoka in Kandahar, Afghanistan was written in Aramaic.
@Grey: Hunters know how to handle animals. They would make good hired help to manage herds. Problem is, if climate is favorable, herds grow over time and need to be moved around because they eat all the fodder.
The herders meanwhile get fat and happy eating all that meat and have lots of babies. These grow up and organize to help manage the growing herds and defend against predators (animal and human). Rinse, repeat, and you have a self-propelled mechanism for an expanding population.
As for language, these herders started out as hired help and part time raiders. They would have learned the "rich folks'" (farmers') language as a matter of practicality (maybe with some nonstandard or "rustic" features). So they would have at least been bi-lingual.
Some might have even spoken a pidgin with simplified grammar rules and mixed vocabulary. "Proper" languages build up a lot of random and arbitrary conventions over time that children and non-native speakers have trouble with - their "beginner's errors" have a tendency to streamline language (paradoxically).
From what part of Finland are your ancestors, Kristiina?
@About Time
"They would make good hired help to manage herds."
Yes
Aboriginal cowboys in Australia
http://resources3.news.com.au/images/2011/06/15/1226075/553655-180611-raparapa.jpg
American Indian cowboys in Florida
http://floridamemory.com/fpc/reference/c013676.jpg
I think the same thing happened in neolithic europe (minus the horses) with the big difference that in America and Australia the farmer's expansion only took a few hundred years which was too fast for the native HGs to adapt into herders completely whereas the european HGs had much longer to adapt.
My mother seems to be nearly totally from Southwest Finland but not from the coast and there seems to be no Swedish speakers among her relatives. I inherited from her a very weird mtDNA line, I5a. I really do not know the story behind it. The places are in Satakunta-Häme: Hämeenkyrö, Mouhijärvi, Sastamala-Vammala. My father has a more mixed origin. His father's line is again from (South)Western Finland, from Ikaalinen, but his yDNA line has been detected also in Ilmajoki area in Bothnia area (Eastern European branch of N1c). My paternal grandmother had the mtDNA line U4d which is concentrated in Southeast Finland, in Iitti-Valkeala area. Her father came from Laukaa area, so he might have represented this Savo expansion.
It seems that all Finns have quite similar amounts of EEF, WHG and ANE, which is of course not a surprise.
When I did Eurogenes K13 admixture test, it was fascinating to have again 1,29% of Amerindian and 1,1% Oceanian. Of course, I had also small percentages of South Asian, East Mediterranean and Northeast African. However, the main components were North Atlantic and Baltic, and after that came Western Mediterranean and Siberian components. I find all this utterly exciting. :-)
Davidski, am I right that your test does not measure ENA? Where do you put the Siberian component in your admixture test with respect to ENA and ANE? My EEF, WHG and ANE percentages amount to 100%. If I have 5.6% of Siberian, do you consider that as ANE or is it just omitted in your test together with for example the Oceanian component?
Siberian and other ENA admixture goes into WHG and ANE, thereby inflating their percentages. Same thing with Oceanian. On the other hand, Sub-Saharan ancestry probably inflates EEF.
But I'm not sure how that works exactly? It probably varies depending on the individual user's K13 results.
EEF 35,39691437
WHG 46,12887156
ANE 18,47421407
I am from Northern Ostrobothnia, but my admix results usually fits poorly to what people think about it, so I usually say Central Ostrobothnia and people are happy with it. My ancestors have however lived here from the 14th centiry or longer. The only study giving comparable results with me is done in Oulu Uniersity by tri Evelina Jakkula.
SpagetiMeatball, I thought you were Italian based on your name.
There is ethnic conflict everywhere and it has always existed. Even in America a country that has nothing to do with genetics and has a young history. Back in colonial times, 1800's, and even into the 1900's there were ethnic conflicts between immigrant groups like Irish and Italians. There used to be ethnic towns(and still are some) where they would speak their native language. Still today you have Italian mafia's which have a lot to do with ethnicity. Inter marriage was looked down on, but after a while children of for example Germans and Swedes were apart of the same culture and mixed, now almost everyone is a mutt.
There has been fighting between white and black, and other forms of racial fighting. There is serious fighting between different towns or neighborhoods, schools(a lot of rivalry is based on sports) everywhere in America. This is totally normal and probably exists in every part of the world. Honestly it can be very fun.
I just commented in the La Brana 1 post that rumour has it that he belonged to C-V20, but no one will probably see it there, so I'm repeating it here. This is the source...
http://www.eupedia.com/forum/threads/29344-Guess-the-Y-haplogroup%28s%29-of-Mesolithic-Iberians-%28Bra%C3%B1a-1-amp-2%29?p=424577&viewfull=1#post424577
My IP address was banned from viewing the Eupedia forum(by Maciamo), so I can't view that. Do you have any idea where this rumor began? I have heard C6 V20 is only in Europe today and has been hypothesized as descended from upper Palaeolithic Europeans like Y DNA I and F 96.
Most of C's clades are in east Eurasians(including Oceania). According to Wikpedia there is C-M356 is in central asia, south asia, and southwest asia, and it also mentioned some possible C*'s in Lebanese. It is surprising early Europeans from the same stock as Loschbour would have a subclade of Y DNA C since they were west Eurasians. IJ, G, and C now seem to be the most likely Y DNA haplogroups of early west Eurasians. I still think R and its father P where originally east Eurasian.
Y DNA C6-V20 and I2a1b*(or intermediate between I2a1 and I2a1b) being native to Mesolithic west Europe is evidence of what I was saying before. Most of the WHG in modern west Europeans is from Mesolithic east Europe because of Indo Europeans. Obviously their paternal lineages didn't survive well. Maybe most of the U5, U2e, and U4 in west Europe today descends from them though. Y DNA I2a2 and I1 may very well be native Mesolithic lineages of central-west Europe and are pretty popular.
R1b in all of Europe was probably in the near east 10,000 years ago. It may make a small affect on total ancestry but R1b L11 alone is evidence Indo Europeans where in the near east some 6,000 years ago.
A new user with the initials CLF, a Spanish flag, and apparently an e-mail address that looks like it's from the lab where the La Brana genomes were tested, basically posted the info at the Eupedia forum. I'm sceptical that it's really Carles Lalueza-Fox, but V20 isn't totally improbable, so I guess we'll see.
By the way, here's the lab website...
http://www.ibe.upf-csic.es/research/research-labs/lalueza-fox.html
Any news on when paper on La Brana-1's is coming out?
I have no idea, and the mysterious CLF didn't say at Eupedia, so we just have to wait.
It was Carles Lalueza-Fox who claimed La Brana-1 had the same gene for blue eyes as modern north Europeans with blue eyes in early December. He was also involved in the first study in 2012. Probably very few people actually know who he is, and would be stupid to think many people would be fooled by having a username with his initials. English is being spoken by people around the world, and if Carles doesn't know it he can use some type of online translate. I think it is him, but its true we will have to wait and see if this is a lie.
It will be interesting to see how La Brana-1 compares to Loschbour and all those other hunter gatherers and farmers. His results in different admixtures seem to say he had some farmer ancestry, like the Gotland hunter gatherers, and unlike Loschbour and the Motolas. The farmer admixture must have been a few generations back since he had vast majority hunter gatherer ancestry, mtDNA U5b2c2, and maybe Y DNA C6-V20. La Brana-1 could probably be put into the EEF, WHG, ANE admixture. I think he will be evidence there is little west European Mesolithic ancestry left, and that most of the WHG in west Europe is from Indo Europeans.
@Barakobama
R1b migrated to the Near East. It isn't from there. R is ANE not EEF. If R1b was in the Near East, then the EEF's would have ANE, which they do not. R1b-L11 is not Near Eastern!!
Davidski,
Could this C group in Western Europe be the reason we see this Oceania, or any other more typically ENA element pop up in Western Europeans? Instead of flow into Europe from more East Eurasia, perhaps? Or is it all just noise, or via third party, ANE (Malta)? I am wondering as you see the English scoring as high as some Siberians in this category, and more than the East Finnish. Just curious as C is important to the Pacific Islands and Western Europe has the highest Oceania of Europeans and is where C is found in Europe. On another note, do you think it is possible that yDNA J is the reason for HG like ancestry in the EEF? Sorry, that is just my interpretation of your comment regarding J sticking around the Near East longer. Thanks.
By other ENA, I mean Siberian, Amerindian(doesn't Loschbour show some at certain K levels?), South East Asian, and Oceania.
CLF, said ten days for the paper. I assume this means, Monday, January 27th.
I just made a thread to spread the rumor.
http://www.theapricity.com/forum/showthread.php?110804-Rearcher-posted-at-Eupedia-La-Brana-1-had-Y-DNA-C-V20
I did a little research and interestingly C-V20 was discovered in southern Europe, and I am not sure if it has been found anywhere else. I would bet it was found specifically in Spain. I think C may have arrived in Europe possibly 40,000 years ago from early east Eurasians not west Eurasians, because besides C-V20 and possibly some other very rare C lineages(learned from Wikpedia) all C lineages are in east Eurasians. Who knows C may have come to Europe some 70,000 years ago and before east and west Eurasian split.
Chad can you please post quotes from what CLF says about La Brana-1? Because my IP address is banned from viewing the forum. Just like I had to do with Laz 2013 I am only going off what people who read the soruce say.
Chad Rohlfsen,
I think J the brotherclade to I which has been found in Mesolithic Luxemburg and Sweden, originated in the near east. J has left ancestry related to what was in WHG there is no doubt about that. So I think what you saying about it giving near easterns some WHG in EEF, WHG, ANE test that is not real WHG is definitely possible.
Y DNA C is so rare in west Europe the population it originated in may have not left a very small amount of blood in modern Europeans. I think your hypothesis could be true though. C is so exclusively east Eurasian(ENA is a lot shorter I will use it for now on) that I doubt it is descended from the same west Eurasians that brought Y DNA I, mtDNA U5, U2e, and U4 and who take up the vast majority of ancestry of Mesolithic Europeans.
All the Mesolithic hunter gatherers from Laz 2013 scored some east Eurasian from K=3-K=9. The Swedish hunter gatherers score some till K=16. It becomes specifically native American green in K=5.There is a tiny tiny pit of yellow east Asian in Loschbour in K=6=K=9. In some of the K's there is dark purple Oceania in all the Mesolithic hunter gatherers. In K=13 Loschbour has some light blue Onge.
Maybe C-V20 is connected to ENA ancestry in Europe.
Is there something particular in your admixture results? I think that my results are not so unexpected. I did both Eurogenes K13 and Dodecad V3 tests, and I see that their groupings are different. The basis components seem to be well in line with normal percentages of people in the Baltic Sea area. On the basis of my previous discussion with Davidski, the trace amounts of Oceanian and Amerindian are typical of Baltic people and Russians and also Finns (?).
When I compare my results with these Finns (https://docs.google.com/spreadsheet/ccc?key=0ArJDEoCgzRKedGR2ZWRoQ0VaWTc0dlV1cHh4ZUNJRUE&pli=1#gid=24), I score a lower Southwest Asian percentage and my Southeast Asian percentage is even 1.22 and my South Asian percentage is also 1.24, while these seem to absent in most Finns. Do not know what I should make of it. :-)
The traces of Oceanian admixture in Europe are just noise, probably caused by shared ancient Eurasian ancestry between Europeans and Oceanians.
And I don't really know what the WHG-like component in EEF represents, and it seems to me that neither do the researchers at this point. It might be something native to the Near East linked to hg J, and/or hunter-gatherer input from southern Europe. Although it might actually represent slightly different things in different people, like Balkan hunter-gatherer ancestry among Northern Europeans, but just Near Eastern admixture among some Southeastern Europeans.
I think it is a possibility C-V20 is connected to ENA ancestry in Europe since C is so non west Eurasian today. I think WHG ancestry in Stuttgart could be mixing with Balkan or central European hunter gatherers. I don't get what your trying to say in the last sentence.
What I'm saying is that EEF might not be exactly the same component across all of West Eurasia or even Europe. For Northern Europeans it might mean ancestry from the first migrant Near Eastern farmers who admixed with the hunter-gatherers in the Balkans. But for Anatolian Turks it might not mean anything but Near Eastern ancestry, including a component native to the Near East that is similar and/or ancestral to WHG, and linked to haplogroups IJ, J and Near Eastern I.
And I don't think C-V20 is an ENA-specific marker. I doubt ENA actually existed when C-V20, or its ancestral clade, moved into Europe. If it did exist, it was probably restricted to a very small part of East Asia. Think about it, there was no ENA in Siberia 24,000 or even 17,000 years ago, and then suddenly it became the most dominant component across East Eurasia.
ENA specific ancestry is old there is no doubt about that maybe some 60,000 years old. It was just that there were more west Eurasian like people that lived in Siberia 17,000-24,000 years ago that doesn't mean there were no ENA people in Siberia or that they didn't exist anywhere at that time.
Pretty much all other C lineages are only in ENA people so it is definitely possible a ENA people brought C-V20 or its ancestral form to Europe. Maybe there was a human migration into Europe that was from over 60,000 years ago and before the ENA and WE(west Eurasian) split. Maybe there are very rare mtDNA haplogroups labeled N*, R*, and M* that descend from these people, probably not.
Laz 2013 I am pretty sure says EEF is mainly descended from something related to what is in the near east today with a little bit of WHG. I don't understand though in K=2-K=20 there isn't one K with a near eastern component. It is still hard to believe there is one ancient component that west Asian, southwest Asian, and Meditreaen mainly descend from.
It makes total sense that most EEF in Europe has a lot of WHG and in the near east is just non WHG west Eurasian ancestry. You think Y DNA I was originally near eastern? I have heard about I* and I2(-I2a) in the near east but I still think it probably originated in Europe. There is probably similar ancestry to WHG in southwest Asian component since it's distribution is similar to Y DNA J1.
Onur, you're like the lone ranger. You appear out of nowhere all guns blazing whenever someone mentions possible South Asian admixture in the Near East.
The South Asian ancestry which I say is almost non-existent in non-Iranic West Asians is ASI ancestry, not ANI ancestry or something similar. Thus it is specifically non-Caucasoid South Asian ancestry which is almost non-existent in non-Iranic West Asians.
Look at the K=16 ADMIXTURE run of Lazaridis et al.:
http://img838.imageshack.us/img838/4694/6pj3.png
As you see, the light green component, which peaks in the Mala, is almost non-existent in West Asia and the Caucasus except in Iranians (unfortunately Kurds and Azeris are not included in that ADMIXTURE analysis, but extant ADMIXTURE analyses suggest that they would have the light green component in levels lower than Iranians but still in non-negligible levels). Looking at its worldwide distribution and levels, it is clear that the light green component is a majority ASI, minority ANI composite component. Georgians, who are a better ANI proxy than are Iranians according to Moorjani et al. 2013, lack that component, which is consistent with the fact that the ANI element of that component is in minority level in its composition while its ASI element is in the majority level.
He's Turkish (as I am).... and naturally we become hectic when somebody mentions some kind of admixture into Turks whether african or indian or whatever.
According to some Kemalist authors, Turks are the pure descendants of the original seljuk nomads who entered anatolia circa 1,200 years ago.
We get touchy about this subject.
I am not a supporter of the idea that Turks are the pure descendants of the original Turkic nomads who entered Anatolia circa 1,000 years ago (it was circa 1,000 years ago, not 1,200 years ago), and I am not touchy about any kind of non-Turkic admixture in Turks. In fact, I acknowledge that Turks are largely of pre-Seljuq Anatolian and/or Balkan ancestry. Also, I am not under the influence of any ideology; I am an open-minded person.
Kristiina
I have same thought that you about the sami admix in finns.
I believe that the reason that Finns is outliers from both balts, russians and scandinavians in these different projects is that there is a large portion of sami mix. To a part it's also because of genetical drift because the population has been very small back in time but the main reason is the sami admix.
"Pretty much all other C lineages are only in ENA people"
Not really true. Australians have yDNA C4, but they do not share the same ancestry with mainland East Asians. C2 is found in Guinea-Oceania, and they also have their own local component which is not the mainland Eurasian East Asian component. C3b is found in America and there is hardly any East Asian in these populations. Greenlanders have C3 without any East Asian at all. Koryaks are mainly C3 and C3c with 0.3% of East Asian. There is C5 in India, but the East Asian ancestry in India peaks with O2 people in Eastern India and not with C5.
Your statement may be true only if we understand ENA as the basic Eastern admixture component seen at level K3, but in this case it is almost as old as Out of Africa and is a very vague concept.
Kristiina
No, my result is typical for Finns in general. But the particular thing appears when we look at the Finnish academic studies where Northern Ostrobothnia is considered by all studies from the Helsinki University to belong to the most Eastern regions in Finland. Just to remember, I am researching history, not my family. I research my family as a genealogist, not genetically. Even this is not something worth to mention if we are not interested in the prehistory of Finland. If we are, then all signals tell us momentous things because small genetic signals are in Finland persistent and revealing, and they are revealing because in Finland we have detectable differences in genetic statistics. So individual results connected to the geography are interesting. You can learn more of this. For example when I pick my 10 closest cousin matches at Gedmatch and compare my ranking on their lists the average ranking is around 50 (rankings 2 to 94 where the second one is the only being my genealogical cousin - proved). Genetic signals tell us about the genetic structure and history which is unquestionable.
Kristiina,
Oceanians, Australians and even Amerindians for the most part are ENA in this paper.
Mickel
Yes, you are right about the Saami admix. But this is not any more a question. The question is where did the Finns come from. It is not possible that we have lived here waiting for Saami people to admix with them.
CLF said:
Yesterday 17:22
sorry, so far everyone is wrong. You have 10 more days for guessing the right answer...
I agree that it makes sense from the European point-of-view to lump a third of world's genetic variation in one "container", because you cannot analyze everything in detail. East Asia and India have been colonized for a very long time, so this ENA does not tell anything about the genetic history of people in these areas. There must have been many waves and migrations in the course of at least 60 000 years of history, and we might find several interesting components there as well.
If this La Braña sequence belongs to yDNA C, it really gives a new perspective to La Braña autosomal results.
Davidski,
That is kind of what I am hinting at, sorry for the poor use of words. What I am wondering is if haplogroup C is not already ENA, if may represent the ancestral population that mixed into Europeans, as well as Siberians, South Asians, Amerindians, and Pacific Islanders. Perhaps Oceania shows up in Europe, for possibly the same reason Amerindian does. An ancestral population that was involved in the creation of another population. Malta, AG1 give Europe the Amerindian, and C-V20 brings Oceanic or any other ENA. Both populations doing the same thing to Europe, but C doing it several thousands of years earlier.
Europe is looking like the drain that sucked in everything but the kitchen sink. I wouldn't bet against there being a fourth and even fifth ancestral population for Europe.
Yes, that's pretty much it. These are just shared layers of ancient Eurasian ancestry being expressed as admixtures from modern populations, because ancient samples aren't available.
But according to the paper, most Europeans only really have two ancestral populations: hunter and farmer. The hunter can be broken down into WHG and ANE, but doesn't need to be for most of the European populations, particularly the mainstream, Indo-European speaking ones.
Some groups from the extremes of the continent do show other influences, like ENA, more recent EEF-like admixture from the Near East, and/or SSA, but they're exceptions. On the other hand, a few isolates don't fit the mainstream hunter (WHG/ANE) and farmer model, and are better fit as hunter (WHG) and farmer, such as the Basques and Sardinians, but again, they're exceptions.
Could it be that HG people are shown to be relatively pure due to high inbreeding? Perhaps they are many different components in one. Perhaps a majority base sharing with ANE, but WHG have a WHG like Near Eastern component, a (pre)ENA C and Amerindian, while ANE are more of he core plus an Amerindian, and maybe a very minor (pre)ENA C. Maybe one component (HG) includes 1,2 or 3 more separate populations.
When I say Amerindian, of course again I mean a population that was involved in creation of Amerindians.
Could there also be an SSA component in UP Europe?
I'm not sure whether most of the modern components we see in various tests existed 24,000 years ago, or even 8,000 years ago.
For instance, the component that Amerindians share with Europeans is ANE. But because MA1 is the only ANE sample in the PCA and ADMIXTURE analyses, he basically comes out like a mix of West Eurasians and Amerindians.
Add another 25 Siberians from around 24,000 years ago to the various tests, and it's very unlikely they'll show any ancestry from modern populations.
"Europe is looking like the drain that sucked in everything but the kitchen sink. I wouldn't bet against there being a fourth and even fifth ancestral population for Europe."
Or there was more than one route out of Africa with various waves passing through Europe often being mostly replaced by later waves leading to a lot of seeming admixture being actually shared ancestry.
1) Atlantic coast -> Iberia
2) East African coast -> Arabia
3) Nile -> Egypt -> Near East
(with 3 being the largest)
Didn't Loschbour and the EEF have an Amerindian related component at higher k levels?
At K=5 level Loschbour, both Motala samples and Mal'ta share the green component with Amerinds but Stuttgart does not! Loscbour has a smaller amount compared to Motala, and Motala samples have c. a third part of the green component of Mal'ta. Also Saami and Chuvash (c. 10%) share a similar amount of green component as Motala samples. Uralic people, Lithuanians, Russians, Ukrainians and Tajiks share a bit smaller amount of green color than Saami and Chuvash. Eskimos and Chukchi have almost 50% of green color. It is also found in Selkup (c. 22%), Nganasan, Dolgan, Yakut, Tuvinian, Altaian, Kirgiz, Kalmyk, Oroqen, Daur, Hezhen, Hazara, Uygur, Uzbek, Turkmen, Nogai, Burusho, Kalash in decreasing order. This pattern is maintained up to K=9 level.
Kristiina,
Is there a 'Papuan' related component in the whg at higher k's?
I wonder how close Papuans would be to West Eurasians, minus the Denisovan. Would the difference be negligible?
@barak
he (CLF) later seems to imply C-V20
R1b is definitely Near Eastern But who says it has to be from the Western parts of the Near East where EEF dominated? R1b is most diverse in the Eastern parts of the Near East, Eastern Anatolia, Iranian plateau, Mesopotamia, the South Caucasus and parts of Central Asia.
Davidski good observation He is obsessed in proving more ASI genes in Kurds and Iranians than there really is. Though Palisto pretty much proved that ASI in Kurds is on average 1% and weaker than in allot of other Near Eastern groups (Lebanese, Syrians, Turks etc) he doesn't seem to accept that. I am getting more and more sure that this guy is actually the Onur who is mostly of Bulgarian Turkish descend and wandering through allot of Anthro-Forums trying to spread his obsession.
ASI is related to Caucasus-Gedrosia and North European anyways so I quite don't get his obsession.
Post a Comment